Valid Names Results
Planococcus citri (Risso, 1813) (Pseudococcidae: Planococcus)Nomenclatural History
- Dorthesia citri Risso 1813: 416. Type data: FRANCE: Menton, on Citrus spp.. Syntypes, female, Type depository: Pretoria: South African National Collection of Insects, South Africa; accepted valid name Notes: This species was described by Risso (1813) as an Ortheziid, but was ultimately placed in the Pseudococcid genus Planococcus by Ferris (1950b). Type material probably lost (Ben-Dov, 1994).
- Coccus tuliparum Bouche 1844: 301. Type data: ENGLAND: London, on Tulipa sp.. Neotype, female, by subsequent designation (BenDovCo1990,81). Type depository: London: The Natural History Museum, England, UK; junior synonym (discovered by Lindin1932f, 201).
- Coccus citri (Risso, 1813); Boisduval 1867: 348. change of combination
- Coccus citry (Risso, 1813); Alfonso 1875: 428. misspelling of species epithet
- Dactylopius citri (Risso, 1813); Signoret 1875c: 312. change of combination
- Dactylopius tuliparum (Bouche, 1844); Signoret 1875c: 323. change of combination
- Dactylopius alaterni Signoret 1875c: 309. Type data: FRANCE: probably Montpellier, on Rhamnus alaternus. Syntypes, female, Type depository: Auckland: New Zealand Arthropod Collection, Landcare Research, New Zealand; Vienna: Naturhistorisches Museum Wien, Austria; junior synonym (discovered by Balach1928c, 143). Notes: Synonymy also by Ben-Dov & Matile-Ferrero (1995:246).
- Dactylopius ceratoniae Signoret 1875c: 311. Type data: FRANCE: Alpes-Maritimes, along the 'Corniche', in bark crevices of Ceratonia siliqua. Syntypes, female, junior synonym (discovered by BenDovMa1995, 246). Notes: Type material probably lost according to Ben-Dov & Matile-Ferrero (1995).
- Dactylopius cyperi Signoret 1875c: 314. Type data: FRANCE: Alpes-Maritimes, Nice, in greenhouse, on Cyperus papyrus. Lectotype, female, by subsequent designation (BenDovMa1995,246). Type depository: Vienna: Naturhistorisches Museum Wien, Austria; junior synonym (discovered by BenDovMa1995, 246). Illustr.
- Dactylopius robiniae Signoret 1875c: 322. Type data: FRANCE: Var, Hyeres and Alpes-Maritimes, Cannes, Nice and Menton on Robinia psedoacacia. Syntypes, female, Type depository: Vienna: Naturhistorisches Museum Wien, Austria; junior synonym (discovered by BenDovMa1995, 249).
- Dactylopius citri; Signoret 1875c: 312. subsequent use Notes: Incorrect citation of author.
- Lecanium phyllococcus Ashmead 1879: 160. Type data: U.S.A.: Florida, on orange trees. Syntypes, female, junior synonym (discovered by Cocker1894, 31). Notes: Type material depository unknown (Cox, 1989).
- Dactylopius brevispinus Targioni Tozzetti 1881: 137. Type data: ITALY: Location not indicated, on various plants in the field. Syntypes, female, junior synonym (discovered by King1900b, 215). Notes: Type material probably lost according to G. Pellizzari-Scaltriti, 1990 personal communication to Y. Ben-Dov.
- Coccus citry (Risso, 1813); Targioni Tozzetti 1881: 134. misspelling of species epithet
- Dactylopius destructor Comstock 1881a: 342. Type data: U.S.A.: on houseplants in greenhouses. Syntypes, female, junior synonym (discovered by Cocker1894, 31). Notes: Type material probably lost (Ben-Dov, 1994).
- Dactylopius farinosus; Cockerell 1898g: 109. misidentification
- Dactylopius secretus Hempel 1900a: 387. Type data: BRAZIL: Ypiranga, in galls on leaves of a plant of the Solanaceae. Syntypes, female, Type depository: Curitiba: Departamento de Zoologia, Setor de Ciencias Biologicas, Universidade Federal do Parana, Brazil; junior synonym (discovered by WilliaGr1992, 394).
- Phenacoccus spiriferus Hempel 1900a: 389. Type data: BRAZIL: Sao Paulo, on cultivated tree. Lectotype, female, by subsequent designation (WilliaGr1992,394). Type depository: Sao Paulo: Museu de Zoologia, Universidade de Sao Paulo, Brazil; junior synonym (discovered by Cox1989, 16).
- Phenacoccus spiniferus Hempel, 1900; Hempel 1901: 110. misspelling of species epithet
- Pseudococcus citri (Risso, 1813); Cockerell 1902p: 252. change of combination
- Dactylopius achado Carus & Field 1903: 230. lapsus calami (discovered by Lindin1936, 286). Notes: misunderstanding of Cockerell (1902) description of Dactylopius citri.
- Pseudococcus alaterni (Signoret, 1875); Fernald 1903b: 97. change of combination
- Pseudococcus ceratoniae (Signoret, 1875); Fernald 1903b: 99. change of combination
- Pseudococcus cyperi (Signoret, 1875); Fernald 1903b: 101. change of combination
- Pseudococcus robiniae (Signoret, 1875); Fernald 1903b: 108. change of combination
- Pseudococcus tuliparum (Bouche, 1844); Fernald 1903b: 111. change of combination
- Dactylopius (Trechocorys) citri (Risso, 1813); Newstead 1908: 9. change of combination
- Pseudococcus citri coleorum Marchal 1908: 236. Type data: FRANCE: Paris, Luxembourg Gardens. Syntypes, female, Type depository: Paris: Museum National d'Histoire naturelle, France; junior synonym (discovered by Borchs1949, 132). Illustr.
- Pseudococcus citri phenacocciformis Brain 1915: 116. Type data: SOUTH AFRICA: Cape Province, Rosebank, on Bouvardia sp.. Syntypes, female, Type depository: Pretoria: South African National Collection of Insects, South Africa; junior synonym (discovered by EzzatMc1956, 65). Illustr.
- Pseudo-Coccus citris Gomez-Menor Ortega 1929: 5. misspelling of both genus and species names
- Planococcus citri (Risso, 1813); Ferris 1950b: 165. change of combination
- Planococcoides cubanensis Ezzat & McConnell 1956: 53. Type data: CUBA: intercepted in U.S.A., on Ficus panduratae. Holotype, female, Type depository: Washington: United States National Entomological Collection, U.S. National Museum of Natural History, District of Columbia, USA; junior synonym (discovered by Cox1989, 16). Illustr.
- Planococcus citricus Ezzat & McConnell 1956: 69. Type data: ITALY: intercepted in U.S.A., on Citrus limonia. Holotype, female, Type depository: Washington: United States National Entomological Collection, U.S. National Museum of Natural History, District of Columbia, USA; junior synonym (discovered by Cox1981, 48). Illustr.
- Planococcus cucurbitae Ezzat & McConnell 1956: 71. Type data: GRENADA: on Cucurbita sp.. Holotype, female, Type depository: Washington: United States National Entomological Collection, U.S. National Museum of Natural History, District of Columbia, USA; junior synonym (discovered by Cox1989, 16). Illustr.
- Planacoccus citri; Meurgey 2011: 82. misspelling of genus name
- Planococcus citi (Risso, 1813); Fallahzadeh & Japoshvili 2017: 34. misspelling of species epithet
- Planoccocus citri Risso Ahmed, et al. 2023: 743. misspelling of both genus and species names
Common Names
- chanchito blanco de los citrus Gonzal2011
- citrus mealybug Essig1915a McKenz1967 SmithBeBr1997
- Citrus mealybug McKenz1967
- cochonilha-algodao CarvalAg1997
- cotonet Lloren1990
- Algodón del Naranjo WatsonKo2022
- citrus wolluis WatsonKo2022
- cocciniglia bianca farinosa della vite WatsonKo2022
- cochenille blanche de l’oranger WatsonKo2022
- common mealybug WatsonKo2022
- Gewaechshausschmierlaus WatsonKo2022
- haknima hakimchit she haadar WatsonKo2022
- kortfrynset uldlus WatsonKo2022
- mikan-kona-kaigaramushi WatsonKo2022
- turuncgil unlu biti WatsonKo2022
- villakilpikirva WatsonKo2022
- بق الموالح الدقيقى AbdRab2012
Ecological Associates
Hosts:
Families: 112 | Genera: 345
- Acanthaceae
- Acanthus | BenDov1994 WilliaGr1992
- Aphelandra squarrosa | AbdulRHe2018
- Blechum pyramidatum | BenDov1994 LincanHoCa2010 WilliaGr1992 | (= Blechnum brownei)
- Eranthemum | BenDov1994 WilliaGr1992
- Fittonia albivenis | BenDov1994
- Hygrophila corymbosa | JansenAl2023 | (=Nomaphila siamensis)
- Justicia carnea | GranarScTe1997
- Pachystachys | BenDov1994
- Pachystachys lutea | YerlikBaKa2023
- Ruellia blechum | BenDov1994 WilliaGr1992
- Adoxaceae
- Viburnum tinus | YerlikBaKa2023 | (= Viburnum lucidum)
- Aizoaceae
- Faucaria tigrina | BenDov2012
- Lithops | JansenAl2023
- Mesembryanthemum cordifolium | YerlikBaKa2023 | (= Aptenia cordifolia)
- Alismataceae
- Alisma | JansenAl2023
- Echinodorus | JansenAl2023
- Amaranthaceae
- Alternanthera echinocephala | LincanHoCa2010
- Alternanthera philoxeroides | BenDov1994
- Amaranthus | BenDov1994
- Beta vulgaris | BenDov2012
- Chenopodium album | AhmedAb2010
- Amaryllidaceae
- Clivia miniata | Suh2020
- Crinum asiaticum | Suh2020 | var. japonicum
- Galanthus elwesii | JansenAl2023
- Hippeastrum | BenDov2012
- Hippeastrum equestre | ZhangDe2023
- Anacardiaceae
- Mangifera indica | BenDov1994 KinjoNaHi1996 McKenz1967 TuWuLe1988 WilliaGr1992 ChowdhRa2024
- Rhus | BenDov1994 WilliaWa1988a
- Schinus molle | KaracaSaSm2020
- Annonaceae
- Annona | Beards1966 BenDov1994
- Annona cherimola | Martin1985 GaimarGoEl2024
- Annona muricata | BenDov1994 LincanHoCa2010 WilliaGr1992
- Annona senegalensis | BenDov1994 Hall1937
- Annona squamosa | BenDov1994 McKenz1967 WilliaGr1992
- Apiaceae
- Apium | BenDov1994 Marott1987a
- Apium graveolens | BenDov2012
- Eryngium foetidum | MirabaGaCa2018
- Hydrocotyle moschata | JansenAl2023 | (=Hydrocotyle sibthorpioides)
- Apocynaceae
- Adenium obesum | Moghad2013
- Asclepias | BenDov2012
- Carissa macrocarpa | AbdulRHe2018 | (= Carissa grandiflora)
- Hoya carnosa | MazzeoSuRu2008
- Hoya nicholsoniae | BenDov1994
- Huernia aspera | LincanHoCa2010
- Mandevilla | JansenAl2023 | (=Dipladenia)
- Nerium oleander | BenDov1994 Martin1985 Suh2020 Tsalev1968 WilliaGr1992 | (= Nerium indicum)
- Rauvolfia verticillata | JansenAl2023
- Stephanotis | BenDov2012
- Tabernaemontana | JansenAl2023
- Vinca minor | JansenAl2023
- Araceae
- Aglaonema robeleynii | BenDov1994 WilliaGr1992 | (= Aglaonema roebelinii)
- Anthurium | BenDov2012
- Anubias barteri | JansenAl2023
- Arisarum vulgare | BenDov1994 Marott1987a
- Cyrtosperma merkusii | BenDov1994 WilliaWa1988a | (= Cryptosperma chamissonis)
- Dieffenbachia | BenDov1994 Marott1987a Osborn1898
- Dieffenbachia amoena | AbdulRHe2018
- Epipremnum aureum | BenDov1994 Marott1987a | (= Scindapsus aureus)
- Lasia spinosa | BugajNJuKa2021
- Philodendron | BenDov1994 WilliaGr1992
- Philodendron hederaceum | AbdulRHe2018 | (= Philodendron scandens)
- Pothos | BenDov2012
- Rhaphidophora | WilliaGr1992
- Spathiphyllum wallisii | AbdulRHe2018
- Syngonium | JansenAl2023
- Syngonium podophyllum | KaracaSa2017
- Araliaceae
- Fatsia japonica | Suh2020
- Hedera rhombea | Suh2020
- Heptapleurum arboricola | GoszczGo2011 | (=Schefflera arboricola)
- Plerandra elegantissima | BenDov1994 Marott1987a | (= Aralia elegantissima) (= Dizygotheca elegantissima)
- Polyscias scutellaria | LincanHoCa2010
- Schefflera | BenDov2012
- Tetrapanax papyrifer | ZhangDe2023
- Arecaceae
- Areca catechu | ZhangDe2023
- Chamaerops | Moghad2013
- Cocos nucifera | BenDov1994 McKenz1967 WilliaGr1992
- Latania | AhmedAb2010
- Livistona chinensis | ZhangDe2023
- Phoenix dactylifera | BenDov1994 McKenz1967
- Saribus rotundifolius | JansenAl2023
- Asparagaceae
- Agave americana | Suh2020
- Asparagus | BenDov1994 McKenz1967 WilliaGr1992
- Asparagus officinalis | LincanHoCa2010
- Asparagus setaceus | ZhangDe2023
- Dracaena | BenDov1994 WilliaGr1992
- Dracaena fragrans | AbdulRHe2018
- Furcraea foetida | LagowsGoHo2022 | (= Furcraea gigantea)
- Ruscus aculeatus | BenDov2012
- Asphodelaceae
- Aloe vera | ZhangDe2023
- Asteraceae
- Artemisia dracunculus | WeintrReMo2017
- Aster | BenDov2012
- Bidens pilosa | BenDov1994 ZhangDe2023 | (= Bidens chilensis)
- Blumea balsamifera | ZhangDe2023
- Conyza bonariensis | YerlikBaKa2023
- Echinops | JansenAl2023
- Erigeron canadensis | ZhangDe2023
- Gerbera | MesselViLe2016
- Helianthus | BenDov2012
- Melanthera biflora | BenDov1994
- Osteospermum ecklonis | YerlikBaKa2023
- Pluchea dioscoridis | EvansAb2023
- Scalesia pedunculata | LincanHoCa2010
- Senecio | BenDov1994 Granar1991
- Solidago | JansenAl2023
- Tagetes | BenDov2012
- Tagetes patula | YerlikBaKa2023
- Xanthium strumarium | BenDov1994 | (= Xanthium occidentale)
- Balsaminaceae
- Impatiens | BenDov1994 McKenz1967
- Impatiens balsamina | YerlikBaKa2023
- Impatiens walleriana | YerlikBaKa2023 MartinClZa2024 | (=Impatiens holstii)
- Begoniaceae
- Begonia | BenDov1994 WilliaGr1992
- Berberidaceae
- Berberis | JansenAl2023
- Nandina domestica | Suh2020
- Bignoniaceae
- Bignonia | BenDov1994 Essig1910c Essig1915a
- Campsis radicans | YerlikBaKa2023
- Jacaranda | BenDov1994
- Tecoma x smithii | AhmedAb2010
- Tecomaria capensis | BenDov1994 Hall1926a | (= Tecoma capensis)
- Boraginaceae
- Cordia dentata | LincanHoCa2010
- Cordia lutea | LincanHoCa2010
- Tournefortia pubescens | LincanHoCa2010
- Trounefortia pubescens | LincanHoCa2010
- Varronia alba | BenDov1994 WilliaGr1992
- Brassicaceae
- Brassica oleracea | BenDov1994 WilliaWa1988a | ssp. capitata
- Cardamine lyrata | JansenAl2023
- Eruca sativa | WeintrReMo2017
- Raphanus sativus | Suh2020
- Bromeliaceae
- Ananas comosus | BenDov1994 McKenz1967
- Burseraceae
- Garuga pinnata | BenDov1994
- Cactaceae
- Astrophytum asterias | JansenAl2023
- Cactus | BenDov1994 McKenz1967
- Cereus | BenDov2012
- Opuntia | BenDov2012
- Opuntia dillenii | ZhangDe2023
- Opuntia ficus-indica | Suh2020
- Campanulaceae
- Campanula pyramidalis | JansenAl2023
- Cannabaceae
- Cannabis sativa | PachecKaBa2020
- Cannaceae
- Canna | YerlikBaKa2023
- Canna coccinea | BenDov1994 TuWuLe1988
- Canna flaccida | BenDov1994 TuWuLe1988
- Canna indica | ZhangDe2023 | (including var. flava)
- Capparaceae
- Capparis spinosa | KaracaSaSm2020
- Caricaceae
- Carica papaya | SosanAjUd2020
- Caryophyllaceae
- Dianthus barbatus | Moghad2013
- Dianthus caryophyllus | AhmedAb2010
- Casuarinaceae
- Casuarina equisetifolia | AhmedAb2010
- Celastraceae
- Euonymus | BenDov2012
- Euonymus japonicus | YerlikBaKa2023
- Cephalotaxaceae
- Cephalotaxus harringtonii | Martin1985 | (= Cephalotaxus drupacea)
- Chloranthaceae
- Ascarina | Willia2013
- Cleomaceae
- Cleome gynandra | EvansAb2023 | (= Gynandropsis pentaphylla)
- Clusiaceae
- Clusia | JansenAl2023
- Garcinia mangostana | Astrid2001
- Combretaceae
- Combretum latifolium | ZhangDe2023
- Commelinaceae
- Tradescantia fluminensis | MdelleAdZo2019
- Convolvulaceae
- Convolvulus arvensis | YerlikBaKa2023
- Ipomoea | BenDov1994
- Ipomoea batatas | BenDov1994 McKenz1967 WilliaGr1992 WilliaWa1988a
- Ipomoea lobata | BenDov1994 | (= Mina lobata)
- Ipomoea pes-caprae | BenDov1994
- Stictocardia tiliifolia | LincanHoCa2010
- Crassulaceae
- Aeonium | PachecKaBa2020
- Crassula | Moghad2013a
- Echeveria | JiSu2012
- Kalanchoe | BenDov2012
- Kalanchoe blossfeldiana | KaracaSaSm2020
- Kalanchoe crenata | PachecKaBa2020 | (= Kalanchoe coccinea)
- Kalanchoe pinnata | ZhangDe2023 | (= Bryophyllum pinnatum)
- Sedum | BenDov2012
- Cucurbitaceae
- Citrullus lanatus | KaracaSaSm2020 | (= Citrullus vulgaris)
- Cucumis melo | BenDov1994 McKenz1967
- Cucumis sativus | JansenAl2023
- Cucurbita maxima | BenDov1994 WilliaWa1988a
- Cucurbita moschata | Daane2000 ZhangDe2023
- Cucurbita pepo | BenDov1994 Essig1910c
- Ecballium elaterium | BenDov2012
- Luffa cylindrica | ZhangDe2023
- Sechium edule | BenDov1994
- Cupressaceae
- Chamaecyparis lawsoniana | Moghad2013a
- Cupressus | Moghad2013
- Cupressus sempervirens | KaracaSaSm2020
- Cycadaceae
- Cycas | AhmedAb2010
- Cyclanthaceae
- Carludovica palmata | ZhangDe2023
- Cyperaceae
- Cyperus | BenDov1994 McKenz1967
- Cyperus alterniflorus | AhmedAb2010
- Cyperus esculentus | GodoiSoMe2024
- Cyperus papyrus | BenDov1994 Cocker1897 Signor1875c
- Didiereaceae
- Portulacaria | JansenAl2023
- Dilleniaceae
- Hibbertia scandens | BenDov1994
- Dioscoreaceae
- Dioscorea | BenDov1994 McKenz1967
- Ebenaceae
- Diospyros | BenDov1994
- Diospyros discolor | ZhangDe2023 | (= Diospyros philippensis)
- Diospyros kaki | BenDov2012
- Ericaceae
- Vaccinium corymbosum | JansenAl2023
- Euphorbiaceae
- Acalypha | BenDov1994 WilliaGr1992
- Codiaeum | BenDov1994 McKenz1967 WilliaGr1992
- Codiaeum variegatum | BenDov1994 Granar1991 MartinLa2011 | (= Croton variegatum)
- Colliguaja odorifera | Mills1984
- Croton | BenDov2012
- Croton scouleri | LincanHoCa2010
- Euphorbia lactea | JansenAl2023
- Euphorbia pulcherrima | BenDov1994 Essig1910c GranarScTe1997
- Jatropha curcas | BenDov1994 WilliaGr1992
- Jatropha podagrica | JansenAl2023
- Macaranga | Beards1966 BenDov1994
- Macaranga tanarius | ZhangDe2023
- Mallotus japonicus | ZhangDe2023
- Manihot esculenta | BenDov1994 Willia1970DJ WilliaGr1992 MurthyChRa2025
- Ricinus communis | LincanHoCa2010
- Fabaceae
- Acacia | KozarGuBa1994
- Acacia confusa | ZhangDe2023
- Albizia julibrissin | YerlikBaKa2023
- Albizia lebbeck | AhmedAb2010
- Arachis hypogaea | BenDov2012
- Archidendron grandiflorum | BenDov1994 | (= Abarema grandiflora)
- Bauhinia | BenDov1994
- Bauhinia purpurea | ZhangDe2023
- Calliandra haematocephala | ZhangDe2023
- Cassia | BenDov1994 WilliaGr1992
- Ceratonia siliqua | BenDov1994 Signor1875c
- Delonix regia | BenDov1994
- Desmodium intortum | MartinLa2011
- Entada phaseoloides | BenDov1994
- Erythrina americana | PadillMaNa2016
- Erythrina caffra | BenDov2012
- Erythrina corallodendron | BenDov1994 WilliaGr1992
- Erythrina variegata | ZhangDe2023
- Falcataria falcata | BenDov1994 WilliaWa1988a | (= Albizia falcataria)
- Gliricidia | BenDov1994 WilliaGr1992
- Glycine max | BenDov1994 WilliaGr1992
- Glycyrrhiza glabra | BenDov2012
- Glycyrrhiza uralensis | YunMoAj2025
- Inocarpus | BenDov1994 WilliaWa1988a
- Lablab purpureus | ZhangDe2023
- Lespedeza | ZhangDe2023
- Leucaena leucocephala | BenDov1994 WilliaWa1988a | (= Leucaena glauca)
- Medicago sativa | BenDov1994
- Mimosa | JansenAl2023
- Neonotonia wightii | BenDov1994 | (= Glycine wightii)
- Phaseolus lunatus | BenDov1994 McKenz1967 | (=Phaseolus limensis)
- Phaseolus vulgaris | BenDov1994
- Pueraria montana | BenDov1994 WilliaWa1988a | (= Pueraria thunbergiana)
- Robinia pseudoacacia | BenDov1994 Signor1875c
- Senna alata | JansenAl2023
- Sophora tomentosa | ZhangDe2023
- Trifolium alexandrinum | BenDov1994 Hall1937
- Trifolium dubium | BenDov1994
- Vigna unguiculata | YerlikBaKa2023 | (= Vigna sinensis)
- Wisteria sinensis | BenDov1994
- Geraniaceae
- Geranium | MifsudMaRu2014
- Pelargonium hortorum | ZhangDe2023
- Pelargonium hybridum | YerlikBaKa2023
- Gesneriaceae
- Saintpaulia ionantha | BenDov2012 | (= Viola africana)
- Goodeniaceae
- Scaevola taccada | BenDov1994 | (= Scaevola koenigii)
- Grossulariaceae
- Ribes | BenDov2012
- Hydrangeaceae
- Hydrangea macrophylla | YerlikBaKa2023
- Iridaceae
- Iris | BenDov2012
- Juglandaceae
- Pterocarya stenoptera | ZhangDe2023
- Lamiaceae
- Ajuga integrifolia | ZhangDe2023 | (= Ajuga bracteosa)
- Clerodendrum | BenDov1994
- Clerodendrum trichotomum | ZhangDe2023
- Coleus | BenDov1994 Cox1987 Essig1915a McKenz1967 WilliaGr1992 BenDov1994 Cox1987 Essig1915a McKenz1967 WilliaGr1992
- Coleus scutellarioides | BenDov1994 Granar1991 | (= Coleus hibridus, Coleus blumei) (= Solenostemon scutellarioides)
- Lamium | JansenAl2023
- Mentha | BenDov1994 WilliaGr1992
- Mentha spicata | KolMaiGhFr2014
- Ocimum | BenDov1994 WilliaWa1988a
- Ocimum basilicum | BenDov1994 WilliaWa1988a
- Physostegia | BenDov2012
- Premna | JansenAl2023
- Salvia rosmarinus | BenDov2012
- Solenostemon | KolMaiGhFr2014
- Tectona grandis | ZhangDe2023 ChowdhRa2024
- Vitex trifolia | BenDov1994
- Lauraceae
- Laurus nobilis | GranarScTe1997
- Persea americana | BenDov1994 McKenz1967 WilliaWa1988a | (= Persea gratissima)
- Lecythidaceae
- Barringtonia | JansenAl2023
- Bertholletia excelsa | BenDov1994
- Lythraceae
- Punica granatum | BenDov1994 Martin1985 McKenz1967
- Magnoliaceae
- Magnolia grandiflora | Suh2020
- Malpighiaceae
- Malpighia glabra | ZhangDe2023
- Malvaceae
- Ceiba pentandra | BenDov1994 WilliaWa1988a
- Commersonia bartramia | BenDov1994
- Durio zibethinus | RahimOh2019 SiriseWaHe2013
- Gossypium hirsutum | BenDov1994 BenDov2012 WilliaGr1992 | (= Gossypium purpurescens)
- Hibiscus | AvasthSh1987 BenDov1994
- Hibiscus mutabilis | Suh2020
- Hibiscus rosa-sinensis | KaracaSaSm2020
- Malva sylvestris | KaracaSaSm2020
- Theobroma cacao | Campbe1983 WilliaGr1992 WilliaWa1988a KoffiBaDe2025
- Tilia tomentosa | YerlikBaKa2023 | (= Tilia argentea)
- Marantaceae
- Ctenanthe | JansenAl2023
- Goeppertia pavonii | ZhangDe2023 | (= Calathea tubispatha)
- Maranta | BenDov1994 Marott1987a
- Thalia | JansenAl2023
- Melastomataceae
- Miconia robinsoniana | LincanHoCa2010
- Menispermaceae
- Legnephora moorii | BenDov1994
- Moraceae
- Artocarpus altilis | LagowsGoHo2022 | (= Artocarpus incisus)
- Artocarpus heterophyllus | AriasCRoHe2022 ChowdhRa2024
- Brosimum utile | BugajNJuKa2021
- Castilloa | BenDov1994 WilliaGr1992
- Ficus | BenDov1994 McKenz1967 Moghad2013
- Ficus altissima | JansenAl2023
- Ficus benjamina | Martin1985 Moghad2013
- Ficus carica | Archan1930 BenDov1994 Bodenh1926 GranarScTe1997 Marott1987a Martin1985
- Ficus elastica | Moghad2013
- Ficus formosana | ZhangDe2023
- Ficus lyrata | PachecKaBa2020
- Ficus microcarpa | ZhangDe2023
- Ficus pandurata | BenDov1994 WilliaGr1992
- Ficus pumila | JansenAl2023
- Ficus sycomorus | AhmedAb2010
- Ficus thonningii | LagowsGoHo2022
- Morus alba | BenDov2012
- Streblus | JansenAl2023
- Musaceae
- Musa | BenDov2012
- Musa acuminata | PerezGCa1987
- Musa paradisiaca | BenDov1994 McKenz1967 WilliaGr1992 | (= Musa sapientum)
- Myrtaceae
- Callistemon lanceolatus | BenDov1994 Essig1910c
- Melaleuca leucadendra | JansenAl2023
- Melaleuca quinquenervia | MilleHeCa2016
- Myrtus communis | BenDov2012
- Psidium guajava | BenDov1994 McKenz1967 WilliaGr1992
- Syzygium | BenDov2012
- Syzygium jambos | JansenAl2023
- Nyctaginaceae
- Bougainvillea | BenDov1994 GranarScTe1997 McKenz1967
- Cryptocarpus pyriformis | LincanHoCa2010
- Oleaceae
- Forsythia intermedia | Moghad2013a
- Fraxinus | WilliaMo2000
- Fraxinus excelsior | Moghad2013
- Jasminum | BenDov1994 WilliaGr1992
- Olea europaea | GranarScTe1997
- Onagraceae
- Fuchsia | BenDov1994 Essig1910c
- Orchidaceae
- Dendrobium | Suh2020
- Myrmecophila tibicinis | BenDov1994 PadillMaNa2016 WilliaGr1992 | (= Schomburgkia tibicinis)
- Prasophyllum | BenDov1994
- Thrixspermum formosanum | ZhangDe2023
- Vanilla africana | BenDov2012
- Orobanchaceae
- Orobanche | EvansAb2023
- Paeoniaceae
- Paeonia | BenDov1994 Essig1910c
- Pandanaceae
- Freycinetia | JansenAl2023
- Pandanus amaryllifolius | ZhangDe2023
- Passifloraceae
- Passiflora edulis | BenDov1994
- Passiflora quadrangularis | GoszczGo2011
- Pentaphylacaceae
- Ternstroemia gymnanthera | Suh2020
- Phyllanthaceae
- Bischofia javanica | ZhangDe2023
- Phyllanthus emblica | SinhaKuKu2021 | (= Emblica officinalis)
- Pittosporaceae
- Pittosporum | BenDov1994 Goux1931
- Pittosporum tobira | Suh2020
- Plantaginaceae
- Veronica | BenDov1994
- Platanaceae
- Platanus orientalis | Archan1930 BenDov1994
- Plumbaginaceae
- Plumbago | BenDov1994 Essig1915a
- Poaceae
- Cynodon dactylon | PellizPoSe2011
- Dendrocalamus hamiltonii | GhoshMe2024
- Echinochloa colonum | AhmedAb2010 | (=Panicum colonum)
- Imperata cylindrica | AhmedAb2010
- Oryza latifolia | BenDov1994 McKenz1967
- Oryza sativa | Moghad2013a
- Saccharum officinarum | BenDov1994
- Polygonaceae
- Persicaria acuminata | BenDov2012 | (= Polygonum acuminatum)
- Polygonum equisetiforme | BenDov2012
- Rumex | LagowsGoHo2022
- Polypodiaceae
- Microsorum | JansenAl2023
- Platycerium bifurcatum | AbdulRHe2018
- Portulacaceae
- Portulaca | MilleHeCa2016
- Portulaca grandiflora | YerlikBaKa2023
- Portulaca oleracea | BenDov1994 WilliaGr1992
- Primulaceae
- Ardisia crenata | Suh2020
- Proteaceae
- Macadamia integrifolia | BenDov1994 Willia1973
- Macadamia tetraphylla | BenDov1994
- Resedaceae
- Caylusea hexagyna | LagowsGoHo2022
- Rhamnaceae
- Ceanothus integerrimus | BenDov1994 Essig1910c
- Rhamnus alaternus | BenDov1994 Cocker1897 Signor1875c
- Sageretia thea | JansenAl2023
- Ziziphus mauritiana | LagowsGoHo2022
- Ziziphus spina-christi | BenDov2012
- Rhizophoraceae
- Kandelia candel | ZhangDe2023 | (= Kandelia rheedei)
- Rosaceae
- Chaenomeles sinensis | Suh2020
- Cydonia oblonga | KaracaSaSm2020
- Fragaria vesca | BenDov1994 McKenz1967
- Malus domestica | BenDov1994 ZhangDe2023 | (= Pyrus malus)
- Malus pumila | GranarScTe1997
- Mespilus germanica | JansenAl2023
- Prunus dulcis | BenDov2012 | (subg. Amygdalus communis)
- Pyrus communis | GranarScTe1997
- Pyrus pyrifolia | Suh2020
- Pyrus ussuriensis | Suh2020
- Rosa | Essig1915a
- Rosa multiflora | Suh2020
- Rosa rugosa | KaracaSaSm2020
- Rubiaceae
- Bouvardia | BenDov1994 Essig1915a
- Coffea | Hall1937
- Coffea arabica | BenDov1994 WilliaGr1992 KozarGuBa1994 RossatAnMa2025
- Coffea canephora | NukmalRoAp2017 | var. robusta
- Coffea liberica | BenDov1994 WilliaGr1992
- Gardenia jasminoides | BenDov1994 Essig1915a GranarScTe1997 Matile1988 | (= Tacsonia jasminoides)
- Ixora chinensis | ZhangDe2023
- Ixora coccinea | LincanHoCa2010
- Manettia schumanniana | BenDov1994 Hall1937
- Morinda umbellata | BenDov1994 WilliaWa1988a | ssp. forsteri)
- Mussaenda erythrophylla | BenDov1994
- Mussaenda shikokiana | ZhangDe2023 | (= Mussaenda esquirolii)
- Neolamarckia cadamba | ZhangDe2023
- Pavetta schumanniana | BenDov1994 Hall1937
- Rutaceae
- Citrus | BenDov1994 CarvalAg1997 Essig1910c KozarGuBa1994 Marott1987a Melis1930 QuezadCoDi1972 UygunSeEr1998
- Citrus aurantiifolia | PadillMaNa2016 SiriseWaHe2013
- Citrus aurantium | BenDov1988b BenDov1994 Bodenh1926 Granar1991 GranarScTe1997 Marott1987a Martin1985 Moghad2013 WilliaGr1992 WilliaWa1988a | (= Citrus bigaradia, Citrus nobilis)
- Citrus hystrix | JansenAl2023
- Citrus junos | Suh2020
- Citrus limon | BenDov1994 GranarScTe1997 Marott1987a Martin1985 WilliaGr1992 | (= Citrus x limonia)
- Citrus maxima | BenDov1994 Essig1910c WilliaWa1988a | (= Citrus decumanus, Citrus grandis)
- Citrus medica | BenDov1994 Essig1910c Moghad2013 | inc. Citrus medica variegata
- Citrus paradisi | KolMaiGhFr2014
- Citrus reticulata | BenDov1994 Granar1991 GranarScTe1997 SiriseWaHe2013
- Citrus sinensis | KolMaiGhFr2014 SiriseWaHe2013
- Citrus unshiu | Suh2020
- Clausena lansium | ZhangDe2023
- Flindersia brayleyana | BenDov1994
- Fortunella | JansenAl2023
- Sapindaceae
- Alectryon tomentosus | BenDov2012
- Dimocarpus longan | MartinLa2011
- Ganophyllum | BenDov1994 BenDov1994
- Nephelium lappaceum | Astrid2001 HernanNiMa2011
- Sapotaceae
- Pouteria campechiana | ZhangDe2023 | (= Lucuma nervosa)
- Saxifragaceae
- Bergenia crassifolia | BenDov1994 Marott1987a
- Tolmiea menziesii | JansenAl2023
- Scrophulariaceae
- Eremophila gilesii | BenDov1994
- Myoporum acuminatum | AhmedAb2010 | (=Myoporum pictum)
- Solanaceae
- Brunfelsia pauciflora | BenDov1994 WilliaGr1992
- Capsicum | PachecKaBa2020
- Capsicum annuum | JansenAl2023
- Cestrum | BenDov1994 Essig1915a
- Datura | LagowsGoHo2022
- Nicotiana tabacum | BenDov1994 McKenz1967
- Salpiglossis | BenDov1994
- Solandra | BenDov1994
- Solanum | BenDov1994 WilliaWa1988a
- Solanum aculeatissimum | ZhangDe2023
- Solanum cheesmaniae | LincanHoCa2010
- Solanum diphyllum | ZhangDe2023
- Solanum douglasii | McKenz1967
- Solanum jasminoides | YerlikBaKa2023
- Solanum lycopersicum | BenDov1994 WilliaGr1992 | (= Lycopersicon esculentum)
- Solanum mauritianum | BenDov1994 WilliaWa1988a | (= Solanum verbascifolium auriculatum)
- Solanum melongena | BenDov1994 McKenz1967
- Solanum muricatum | KaracaSaSm2020
- Solanum nigrum | BenDov2012
- Solanum pseudocapsicum | ZhangDe2023
- Solanum tuberosum | BenDov1994 GranarScTe1997 WilliaGr1992
- Strelitziaceae
- Strelitzia alba | Moghad2013
- Strelitzia nicolai | BenDov2012
- Strelitzia reginae | BenDov1994 Essig1910c Essig1915a | (= Strelitzia gigantea)
- Tamaricaceae
- Tamarix aphylla | BenDov1994 Matile1988
- Theaceae
- Camellia sinensis | ZhangDe2023
- Thymelaeaceae
- Thymelaea hirsuta | BenDov2012
- Urticaceae
- Cecropia | JansenAl2023
- Dendrocnide | JansenAl2023
- Dendrocnide moroides | JansenAl2023
- Parietaria | BenDov2012 Bodenh1926 PellizPoSe2011
- Parietaria judaica | YerlikBaKa2023
- Parietaria officinalis | BenDov1994 Marott1987a
- Verbenaceae
- Duranta | BenDov2012
- Duranta erecta | BugajNJuKa2021
- Lantana camara | ZhangDe2023
- Lippia | AhmedAb2010
- Verbena | ZhangDe2023
- Vitaceae
- Cissus | BenDov2012
- Parthenocissus quinquefolia | YerlikBaKa2023
- Vitis | WilliaMo2000
- Vitis vinifera | BenDov1994 GranarScTe1997 Marott1987a Martin1985 UygunSeEr1998
- Zygophyllaceae
- Balanites aegyptiaca | BenDov2012
- Zygophyllum | EvansAb2023
Foes:
Families: 25 | Genera: 69
- Amaryllidaceae
- Allium sativum | ModaffGiUr2025 | essential oil
- Aphelinidae
- Aphytis alami | WatsonKo2022
- Coccobius fulvus | WatsonKo2022
- Coccophagus | Moore1988
- Coccophagus caridei | Moore1988
- Coccophagus gurneyi | Blumbe1997
- Coccophagus lycimnia | Bolu2012
- Coccophagus pseudococci | WatsonKo2022
- Encarsia udaipuriensis | WatsonKo2022
- Marietta connecta | WatsonKo2022
- Marietta leopardina | WatsonKo2022
- Marietta picta | WatsonKo2022
- Asteraceae
- Chrysanthemum | YerlikBaKa2023
- Bacillaceae
- Bacillus velezensis | YunMoAj2025
- Cecidomyiidae
- Coccodiplosis citri | Barnes1935
- Diadiplosis | AhmedAb2010
- Chamaemyiidae
- Leucopis | GaimarGoEl2024
- Leucopis cortesi | GaimarGoEl2024
- Chrysopidae
- Chrysoperla carnea | NukmalRoAp2017 AhmedAb2010 | (=Chrysopa vulgaris aegyptica)
- Chrysoperla externa | PinheiCoCa2023
- Chrysoperla lucasina | MesselViLe2016
- Plesiochrysa lacciperda | Krishn1988 | (= Chrysopa lacciperda)
- Clavicipitaceae
- Metarhizium anisopliae | KaracaKaSi2016
- Coccinellidae
- Brumoides lineatus | WengHu1988
- Chilocorus bipustulatus | BalalaShSh2025
- Cryptolaemus montrouzieri | BaskarLaUt1999 Clause1978 Fisher1963 JayaraChGo1988 MichelHa1995
- Diomus austrinus | ChongOeOs2005
- Diomus ochroderus | BennetSi1964 | (= Scymnus ochroderus)
- Diomus pumilio | Meyerd1983 | (= Diomas flavifrons)
- Exochomus metallicus | Moore1988
- Hyperaspis | NukmalRoAp2017
- Nephus (Bipunctatus) conjunctus | ArgyriStMo1976 KontodLyKa2005 | (= Nephus (Bipunctatus) includens)
- Nephus (Sidis) hiekei | AlAnsiAlFa2020 ArgyriStMo1976 | (= Scymnus hieki)
- Nephus bipunctatus | Moore1988
- Nephus bisignatus | KontodLyKa2005 MichelHa1995
- Nephus reunioni | HennekOpAl1987 | (= Scymnus reunioni)
- Novius cardinalis | AhmedAb2010 | (= Rodolia cardinalis)
- Rhyzobius lophanthae | NukmalRoAp2017
- Scymnus | ArgyriStMo1976 BennetSi1964 JayaraChGo1988
- Scymnus (Pullus) syoitii | PooranAnTh2024
- Scymnus agrumi | AlAnsiAlFa2020
- Scymnus coccivora | VidyaBh2017
- Scymnus includens | PriesnHo1940
- Scymnus interruptus | AhmedAb2010
- Scymnus pyrocheilus | VidyaBh2017
- Scymnus seriacus | AhmedAb2010
- Scymnus subvillosus | UlgentSzUy2013
- Tenuisvalvae bisquinquepustulata | SantosCaPe2017
- Tenuisvalvae notata | CarmoSiTo2026
- Cordycipitaceae
- Akanthomyces lecanii | ArmandKhNa2024
- Akanthomyces muscarius | ArmandKhNa2024
- Beauveria bassiana | ElAalaRaSb2024a
- Beauveria brongniartii | KaracaKaSi2016
- Isaria farinosa | KaracaKaSi2016
- Lecanicillium lecanii | KaracaKaSi2016
- Davidiellaceae
- Cladosporium oxysporum | Moore2002
- Encyrtidae
- Acerophagus angelicus | WatsonKo2022
- Acerophagus mundus | Clause1978 | (=Pseudaphycus mundus)
- Acerophagus nubilipennis | WatsonKo2022
- Acerophagus perdignus | WatsonKo2022
- Aenasius abengouroui | WatsonKo2022
- Aenasius advena | TangaGhTo2021
- Aenasius arizonensis | JaposhYeKa2023
- Aenasius martinii | WatsonKo2022
- Aenasius masii | WatsonKo2022
- Agarwalencyrtus citri | Trjapi1989
- Alamella flava | WatsonKo2022
- Anagyrus abyssinicus | WatsonKo2022
- Anagyrus agraensis | WatsonKo2022
- Anagyrus amoenus | WatsonKo2022
- Anagyrus ananatis | WatsonKo2022
- Anagyrus aurantifrons | Comper1939d
- Anagyrus beneficians | WatsonKo2022
- Anagyrus bohemanni | Tsalev1972
- Anagyrus chrysos | WatsonKo2022
- Anagyrus coccidivorus | WatsonKo2022
- Anagyrus dactylopii | BasheeAsRa2014
- Anagyrus fusciventris | BlumbeVa2001
- Anagyrus greeni | AbdRab2001d
- Anagyrus kamali | WatsonKo2022
- Anagyrus kivuensis | WatsonKo2022
- Anagyrus mirzai | WatsonKo2022
- Anagyrus nigrescens | WatsonKo2022
- Anagyrus novickyi | MoghadWa2022
- Anagyrus pseudococci | Blumbe1997 Clause1978 MahfouDh2009 Moore1988
- Anagyrus sawadai | WatsonKo2022
- Anagyrus schoenherri | WatsonKo2022
- Anagyrus tibimaculatus | WatsonKo2022
- Anagyrus vladimiri | IzraelLaNe2020 | (= Anagyrus sp. near pseudococci)
- Blepyrus insularis | AbdRab2001d
- Blepyrus saccharicola | WatsonKo2022
- Bothriocraera bicolor | WatsonKo2022
- Cerapterocerus virens | WatsonKo2022
- Cheiloneurus carinatus | WatsonKo2022
- Cheiloneurus claviger | WatsonKo2022
- Cheiloneurus cyanonotus | WatsonKo2022
- Cheiloneurus flaccus | WatsonKo2022
- Cheiloneurus gahani | WatsonKo2022
- Chrysoplatycerus howardii | WatsonKo2022
- Chrysoplatycerus splendens | WatsonKo2022
- Clausenia confusor | WatsonKo2022
- Clausenia guineensis | WatsonKo2022
- Clausenia josefi | WatsonKo2022
- Clausenia purpurea | Suh2019b
- Coccidoctonus pseudococci | WatsonKo2022
- Coccidoctonus trinidadensis | WatsonKo2022
- Coccidoxenoides perminutus | CeballWa2004 Clause1978 Golber1982 JoyceHoBe2001 MahfouDh2009 Moore1988 Trjapi1989 WatsonKo2022 | (= Pauridia peregrina),
- Discodes aeneus | WatsonKo2022
- Encyrtus aurantii | WatsonKo2022
- Gahaniella tertia | WatsonKo2022
- Gyranusoidea flava | WatsonKo2022
- Homalotylus quaylei | Trjapi1989
- Leptomastidea abnormis | ArgyriStMo1976 Clause1978 Flande1953a HennekOpAl1987 MahfouDh2009 Moore1988 Trjapi1989 | (= Leptomastidea aurantiaca)
- Leptomastix dactylopii | BasheeAsRa2014 Blumbe1997 Clause1978 MahfouDh2009 Moore1988 Trjapi1989 | (= Leptomastix bifasciatus)
- Leptomastix epona | BlumbeVa2001
- Leptomastix flava | AbdRab2001d
- Neoplatycerus palestinensis | Rivnay1945 Trjapi1989
- Prochiloneurus aegyptiacus | Rivnay1945 Trjapi1989 | (= Achrysopophagus aegyptiacus)
- Prochiloneurus io | Trjapi1989
- Tropidophryne natalensis | Comper1939d
- Geocoridae
- Geocoris ochropterus | NhuQuCh2025
- Heliodinidae
- Stathmopoda auriferella | AhmedAb2010
- Hemerobiidae
- Sympherobius fallax | MangouAli2009 | (=Sympherobius sanctus)
- Sympherobius pygmaeus | YaylaTuSa2020
- Heterohabditidae
- Heterohabditis bacteriophora | NiekerMa2012
- Heterorhabditidae
- Heterorhabditis bacteriophora | MamukYuCa2024
- Heterorhabditis indica | MamukYuCa2024
- Heterorhabditis safricana | NiekerMa2012
- Heterorhabditis zealandica | DuncanMa2024 NiekerMa2012
- Lycaenidae
- Spalgis epius | DineshVe2016
- Noctuidae
- Autoba beraudi | AhmedAb2010
- Autoba gayneri | AhmedAb2010
- Rivula sericealis | AhmedAb2010
- Platygastridae
- Allotropa kamburovi | AnneckPr1976a
- Pteromalidae
- Pachyneuron muscarum | ArgyriStMo1976 | (= Pachyneuron concolor)
- Salticidae
- Plexippus paykulli | NukmalRoAp2017
- Thyene imperialis | NukmalRoAp2017
- Signiphoridae
- Chartocerus kurdjumovi | JalilvShFa2014
- Steinernematidae
- Steinernema bicornutum | MamukYuCa2024
- Steinernema carpocapsae | MamukYuCa2024
- Steinernema citrae | NiekerMa2012
- Steinernema khoisanae | NiekerMa2012
- Steinernema yirgalemense | DuncanMa2024 NiekerMa2012
- Trombidiidae
- Allothrombium pulvinum | SaboorHoHa2003
Associates:
Families: 10 | Genera: 24
- Betaflexiviridae
- Grapevine virus A (GVA) | HerrbaAlPr2017
- Grapevine virus B (GVB) | HerrbaAlPr2017
- Betaproteobacteria
- Candidatus Tremblaya | GruwelVoPa2005
- Candidatus Tremblaya princeps | ChoiLe2022
- Caulimoviridae
- Banana streak virus (BSV) | PalmaJBlGu2019 PooranAnTh2024
- Cacao Mild Mosaic Virus | PuigWuSu2021
- Cacao bacilliform Sri Lanka virus | WatsonKo2022
- Cacao swollen shoot virus | WatsonKo2022 GyamweDoAk2023 KoffiBaDe2025
- Citrus yellow mosaic virus | AhmedApKa2023
- Dioscorea alata bacilliform virus | WatsonKo2022
- Piper yellow mottle virus (PYMoV) | AhmedApKa2023
- SRV | AhmedApKa2023
- Schefflera ringspot virus | WatsonKo2022
- Closteroviridae
- GLRaV-1 | HerrbaAlPr2017
- GLRaV-3 | BahderPoAl2013
- Enterobacteraceae
- Candidatus Moranella endobia | WangLiRu2023
- Formicidae
- Camponotus | WatsonKo2022
- Camponotus compressus | KavyaYHeBh2026
- Crematogaster | WatsonKo2022
- Crematogaster brunnea | KavyaYHeBh2026 | subsp. rabula
- Crematogaster rogenhoferi | KavyaYHeBh2026
- Dolichoderus taprobanae | KavyaYHeBh2026
- Lasius grandis | PerezPeTe2021 PlataGoBe2023a
- Lasius niger | PerezPeTe2021
- Linepithema humile | MansouMaLa2011
- Monomorium | KavyaYHeBh2026
- Oecophllya smaragdina | BluthgFi2002
- Oecophylla smaragdina | KavyaYHeBh2026
- Paratrechina longicornis | KavyaYHeBh2026
- Pheidole | WatsonKo2022
- Solenopsis geminata | KavyaYHeBh2026
- Solenopsis saevissima | GuindaNoWo2017
- Tapinoma melanocephalum | KavyaYHeBh2026
- Tapinoma nigerrimum | MansouMaLa2011
- Technomyrmex albipes | KavyaYHeBh2026
- Trichomyrmex destructor | KavyaYHeBh2026
- Wasmannia auropunctata | WatsonKo2022
- Meruliaceae
- Diacanthodes novoguineensis | CarmonBeMe2025
- Pseudococcidae
- Phenacoccus miruku | PowellDeMo2024
- Pseudomonadaceae
- Pseudomonas putida | HarishAr2022
- Pyralidae
- Cryptoblabes gnidiella | FrancoZaMe2009 WatsonKo2022
Geographic Distribution
Countries: 176
- Afghanistan | KozarFoZa1996
- Algeria | Balach1927 BenDov1994
- Angola | Almeid1973 BenDov1994 DeLott1967a
- Antigua and Barbuda
- Antigua | BenDov1994 WilliaGr1992
- Argentina | CABI1999
- Buenos Aires | BenDov1994 Granar1991
- Catamarca | GranarCl2003
- Cordoba | BenDov1994 Granar1991
- Entre Rios | GranarCl2003
- Salta | GranarCl2003
- Tucuman | BenDov1994 Granar1979 Granar1991 WilliaGr1992
- Armenia | BenDov1994 TerGri1956 TerGri1966a
- Australia
- Australian Capital Territory | BenDov1994 Willia1985
- New South Wales | BenDov1994 Willia1985
- Northern Territory | BenDov1994 SmithBeBr1997 Willia1985
- Queensland | BenDov1994 SmithBeBr1997 Willia1985
- South Australia | BenDov1994 Willia1985
- Tasmania | BenDov1994 Willia1985
- Western Australia | BenDov1994 SmithBeBr1997 Willia1985
- Austria | Malump2011a
- Azerbaijan | BenDov1994 Terezn1975
- Azores | CarvalFrAg1996 LopesFiMa2008
- Bahamas | WatsonKo2022
- Balearic Islands | CABI1999
- Bangladesh | BenDov1994 Varshn1992
- Barbados | WatsonKo2022
- Belgium | CABI1999
- Benin | CABI1999
- Bermuda | BenDov1994 CABI1999 HodgsoHi1990 HodgsoHi1991 WilliaGr1992
- Bonin Islands (=Ogasawara-Gunto) | Kawai1987 KawaiMaUm1971
- Brazil | BenDov1994 Hempel1900a WilliaGr1992
- Espirito Santo | CulikMaVe2007
- Minas Gerais | CABI1999
- Parana | FoldiKo2006
- Pernambuco | PachecKaSi2019
- Rio Grande do Sul | BertelBa1966 GomesCRe1949
- Rio de Janeiro | CABI1999
- Santa Catarina | CABI1999
- Sao Paulo | CABI1999
- British Virgin Islands | CABI1999
- Brunei | CABI1999
- Bulgaria | BenDov1994 TrenchTrTo2010 Tsalev1968 Tsalev1972 Tschor1939
- Burkina Faso | CABI1999
- Burma (=Myanmar) | CABI1999
- Burundi | CABI1999
- Cameroon | CABI1999
- Canada
- Ontario | McLeod1939
- Canary Islands | MatileOr2001 PerezGCa1987
- Cape Verde | SchmutPiKl1978 VanHarCoWi1990
- Cayman Islands | BenDov1994 WilliaGr1992
- Chad | CABI1999
- Chile | CABI1999
- Atacama | Gonzal2011
- Coquimbo | PizarrAlBa2020
- Metropolitana | CorreaLoMa2015
- O'Higgins | Gonzal2011
- China | BenDov1994 Borchs1956b Tang1992 TangHaTa1992
- Anhui (=Anhwei) | CABI1999
- Beijing (=Peking) | ZhangDe2023
- Fujian (=Fukien) | CABI1999
- Guangdong (=Kwangtung) | CABI1999
- Guangxi (=Kwangsi) | CABI1999
- Guizhou (=Kweichow) | CABI1999
- Hainan | CABI1999
- Hebei (=Hopei) | WangZhTi2018
- Hubei (=Hupei) | CABI1999
- Hunan | BenDov1994 HuHeWa1992
- Jiangsu (=Kiangsu) | CABI1999
- Jiangxi (=Kiangsi) | CABI1999
- Shanghai | ZhangDe2023
- Sichuan (=Szechwan) | CABI1999
- Xianggang (=Hong Kong) | CABI1999
- Xinjiang Uygur (=Sinkiang) | ZhangDe2023
- Yunnan | CABI1999
- Zhejiang (=Chekiang) | CABI1999
- Colombia | BenDov1994 Kondo2001 Kondo2008a KondoRaVe2008 WilliaGr1992
- Comoros | BenDov1994 Matile1978
- Congo | WatsonKo2022
- Cook Islands | BenDov1994 WilliaWa1988a
- Corsica | Foldi2003
- Costa Rica | BenDov1994 WilliaGr1992
- Cote d'Ivoire (=Ivory Coast) | CABI1999
- Crete | Argyri1983 Ayouta1940 BenDov1994 PellizPoSe2011
- Croatia | WatsonKo2022
- Cuba | BenDov1994 WilliaGr1992
- Cyprus | CABI1969
- Czech Republic | BenDov1994 Zahrad1990
- Democratic Republic of the Congo (=Zaire) | WatsonKo2022
- Dominica | BenDov1994 WilliaGr1992
- Dominican Republic | BenDov1994 WilliaGr1992
- Easter Island (=Rapa Nui) | WynneHoCo2025
- Ecuador | BenDov1994 WilliaGr1992
- Egypt | BenDov1994 Hall1926a
- El Salvador | BenDov1994 QuezadCoDi1972
- Eritrea | CABI1999
- Ethiopia | CABI1999
- Federated States of Micronesia | CABI1999
- Caroline Islands | Beards1966 BenDov1994
- Kosrae (=Kusaie) | Beards1966 BenDov1994
- Pohnpei Island | Beards1966 BenDov1994
- Truk Islands | Beards1966 BenDov1994
- Yap | Beards1966 BenDov1994
- Fiji | Thaman2018
- France | BenDov1994 Cocker1897 Foldi2001 Goux1931 MatileGePi2004 Risso1813 Signor1875c
- French Guiana | WatsonKo2022
- French Polynesia | BenDov1994 WilliaWa1988a
- Gabon | CABI1999
- Galapagos Islands | BenDov1994 CaustoPeSi2006 LincanHoCa2010 WilliaGr1992
- Georgia (Republic of) | BenDov1994 Terezn1975 YasnosTaCh2005
- Germany | WatsonKo2022
- Ghana | BenDov1994 Campbe1983
- Greece | Argyri1983 MichelHa1995
- Grenada | BenDov1994 EzzatMc1956 WilliaGr1992
- Guadeloupe | MatileEt2006
- Guam | CABI1999
- Guatemala | BenDov1994 WilliaGr1992
- Guiana | Remill1988
- Guinea | WatsonKo2022
- Guyana (=British Guiana) | BenDov1994 WilliaGr1992
- Haiti | PerezG2008
- Hawaiian Islands | CABI1999
- Maui | Zimmer1948
- Molokai | Zimmer1948
- Oahu | Zimmer1948
- Honduras | BenDov1994 WilliaGr1992
- Hungary | BenDov1994 KosztaKo1978
- India | BenDov1994 Varshn1992
- Gujarat | Willia2004a
- Karnataka | Willia2004a
- Kerala | Willia2004a
- Maharashtra | Willia2004a
- Meghalaya | WatsonKo2022
- Sikkim | Willia2004a
- Tamil Nadu | CABI1999
- Tripura | CABI1999
- Uttar Pradesh | CABI1999
- West Bengal | Nath1972 Willia2004a
- Indonesia | CABI1999
- Java | BenDov1994 Betrem1937 Willia2004a
- Kalimantan (=Borneo) | CABI1999
- Sulawesi (=Celebes) | CABI1999
- Sumatra | Green1930c
- Iran | KozarFoZa1996 Moghad2009 WilliaMo2000
- Iraq | BenDov1994 Bodenh1943
- Ireland | OConnoGeMa2013
- Israel | Avidov1961 BenDov1988b BenDov1994 Bodenh1924
- Italy | BenDov1994 LongoMaPe1995 Marott1987a
- Sicily | RussoMa1997
- Jamaica | BenDov1994 WilliaGr1992
- Japan | KinjoNaHi1996
- Jordan | BenDov2006a
- Kampuchea (=Cambodia) | CABI1999
- Kenya | CABI1999
- Kiribati
- Gilbert Islands | Beards1966 BenDov1994
- Laos | CABI1999
- Lebanon | BenDov1994 Bodenh1926 Talhou1969
- Liberia | CABI1999
- Libya | Martin1954
- Luxembourg | Gertso2016
- Madagascar | CABI1999
- Madeira Islands | CarvalAg1997 CarvalFrAg1996
- Malawi | CABI1999
- Malaysia
- Malaya | Willia2004a
- Sabah | CABI1999
- Sarawak | CABI1999
- Maldives | WatsonOoGi1995 Willia2004a
- Mali | WatsonKo2022
- Malta | Borg1919 MifsudMaRu2014
- Marshall Islands | Beards1966 BenDov1994
- Martinique | MatileEt2006
- Mauritius | CABI1999
- Mexico | BenDov1994 WilliaGr1992
- Morelos | PadillMaNa2016
- Nayarit | ChamLuRo2019
- Nuevo Leon | PadillMaNa2016
- Oaxaca | PadillMaNa2016
- Puebla | PadillMaNa2016
- Montserrat | CABI1999
- Morocco | Rungs1970
- Mozambique | CABI1999
- Netherlands | HennekOpAl1987 MesselViLe2016
- New Caledonia | MilleHeCa2016
- New Zealand | BenDov1994 Cox1987
- Niger | WatsonKo2022
- Nigeria | CABI1999
- Niue | BenDov1994 WilliaWa1988a
- Oman | WatsonKo2022
- Pakistan | BenDov1994 Varshn1992 Willia2004a
- Palau | CABI1999
- Papua New Guinea | BenDov1994 CABI1999 WilliaWa1988a
- Paraguay | BenDov1994 WilliaGr1992
- Peru | Beders1969 BenDov1994 DelaCrCh1996 WilliaGr1992
- Philippines | CABI1999
- Luzon | Willia2004a
- Poland | BenDov1994 Dziedz1990 Koteja1974b
- Portugal | BenDov1994 CarvalFrAg1996 Fernan1992 FrancoPa1991 Seabra1930
- Puerto Rico & Vieques Island
- Puerto Rico | BenDov1994 Martor1976 WilliaGr1992
- Reunion | GermaiMiPa2014 Mamet1957
- Romania | Savesc1960 Teodor2018
- Russia | WatsonKo2022
- Rwanda | CABI1999
- Ryukyu Islands (=Nansei Shoto) | CABI1999
- Saba & Sint Eustatius (=Neth. Leew. Is.)
- Saba | WatsonKo2022
- Saint Croix | Beatty1944
- Saint Helena | CABI1999
- Saint Lucia | Malump2012b
- Saint Vincent and the Grenadines | CABI1999
- San Marino | BenDov1994 WilliaGr1992
- Sao Tome and Principe
- Sao Tome | BenDov1994 Laing1928 Seabra1921
- Sardinia | BenDov1994 Melis1930 PellizFo1996
- Saudi Arabia | Beccar1971 BenDov1994 Matile1984c
- Senegal | CABI1999
- Seychelles | CABI1999
- Aldabra Island | WilliaMa2009b
- Aldabra Island | WatsonKo2022
- Sierra Leone | CABI1999
- Slovenia | Janezi1954 Seljak2010
- South Africa | BenDov1994
- South Korea | ESKSAE1994 KwonDaPa2003a
- Spain | Martin1985 PlataGoBe2023c
- Sri Lanka | BenDov1994 Varshn1992 Willia1999 Willia2004a
- Sudan | CABI1999
- Suriname | BenDov1994 WilliaGr1992
- Swaziland (=Eswatini) | CABI1999
- Switzerland | KozarGuBa1994
- Syria | BasheeAsRa2014 BenDov1994 Talhou1969
- Taiwan | BenDov1994 TuWuLe1988 WongChCh1999
- Tajikistan (=Tadzhikistan) | BenDov1994 Terezn1975
- Tanzania | CABI1999
- Thailand | Willia2004a
- Togo | CABI1999
- Tonga | BenDov1994 WilliaWa1988a
- Trinidad and Tobago | CABI1999
- Tunisia | CABI1969 HalimaGeMd2014 MahfouDh2009
- Turkey | BenDov1994 Bodenh1953a KaydanKiKo2005 Tuncyu1970a UygunSeEr1998
- Turkmenistan | Archan1930 BenDov1994 Terezn1975
- U.S. Virgin Islands | CABI1999
- Uganda | CABI1999
- Ukraine | CABI1999
- Krym (=Crimea) Oblast | BenDov1994 Terezn1975
- United Kingdom | CABI1999
- England | MalumpBa2012
- Scotland | WatsonKo2022
- Wales | CABI1999
- United States
- Alabama | WatsonKo2022
- Arizona | CABI1999
- Arkansas | CABI1999
- California | BenDov1994 Essig1909a Essig1915a Ferris1918d McKenz1967
- Florida | Speare1922
- Iowa | BenDov1994 Osborn1898
- Kansas | CABI1999
- Louisiana | CABI1999
- Maryland | CABI1999
- Massachusetts | CABI1999
- Missouri | CABI1999
- New Mexico | CABI1999
- Ohio | CABI1999
- South Carolina | CABI1999
- Tennessee | CABI1999
- Texas | CABI1999
- Virginia | CABI1999
- Uruguay | GranarScTe1997 PachecKaBa2020
- Uzbekistan | BenDov1994 Terezn1975
- Venezuela | BenDov1994 FoldiKo2006 WilliaGr1992
- Vietnam | BenDov1994 DanzigKo1990 Willia2004a
- Western Samoa | BenDov1994 Laing1927 WilliaWa1988a | WatsonKo2022
- Yemen | CABI1999
- Yugoslavia | Masten2007 MastenSiKr2008
- Zambia | CABI1999
- Zanzibar | WilliaMa2005a
- Zimbabwe | BenDov1994 Hall1937 Hodgso1969c
Keys
- ZarkanHaRe2025: pp.56-58 ( Adult (F) ) [Planococcus in Southeast and southern Asia]
- BeltraLlHu2025: pp.114 ( Adult (F) ) [Scale insects found on avocados, olives, and grapes in Arequipa, Peru]
- AhmedPoMi2025: pp.9 ( Adult (F) ) [Florida Pseudococcidae]
- ZarkanFaAp2024: pp.234-238 ( Adult (F) ) [Planococcus in southern Asia]
- CaballKo2024: pp.9 ( Adult (F) ) [Species associated with coffee roots in Colombia]
- SzitaFeGe2023: pp.5 ( Adult (F) ) [Pseudococcidae on imported fruits in Hungary]
- EvansAb2023: pp.8-13 ( Adult (F) ) [Pseudococccidae of Egypt]
- ZhangDe2023: pp.79-80 ( Adult (F) ) [Chinese Planococcus]
- MoghadWa2022: pp.107 ( Adult (F) ) [species of Planococcus in Iran]
- JoshiSuRe2021: pp.118-119 ( Adult (F) ) [Field characters of mealybugs in cassava]
- JoshiSuRe2021: pp.119-120 ( Adult (F) ) [Microscopic characters of mealybugs in cassava]
- MoghadNe2020: pp.387-388 ( Adult (F) ) [Planococcus in Iran]
- Moghad2020: pp.387-388 ( Adult (F) ) [Planococcus in Iran]
- Hodgso2020: pp.32-34 ( Adult (M) ) [higher groups of Pseudococcidae]
- PachecKaBa2020: pp.504-505 ( Adult (F) ) [Pseudococcidae in Uruguay]
- Suh2019a: pp.2-6 ( Adult (F) ) [mealybugs interepted in S. Korea]
- DanzigGa2015: pp.207 ( Adult (F) ) [Planococcus species]
- MoghadEs2014: pp.59 ( Adult (F) ) [Species on Ficus in Iran]
- Moghad2013: pp.63 ( ) [Key to species of Genus Planococcus Ferris from Iran]
- SiriseWaHe2013: pp.78-80 ( Adult (F) ) [Mealybugs on Fruit Crops in Sri Lanka]
- ParsaKoWi2012: pp.10-Aug ( Adult (F) ) [Key to mealybugs (Hemiptera: Pseudococcidae) recorded on Manihot spp. (Euphorbiaceae) in the World]
- Gonzal2011: pp.135 ( Adult (F) ) [Key to separating species of Planococcus on fruit]
- WilliaMa2009a: pp.97-101 ( Adult (F) ) [Pseudococcidae species from Mauritius]
- Willia2004a: pp.620-622 ( Adult (F) ) [Planococcus species of southern Asia]
- KwonDaPa2003a: pp.406 ( Adult (F) ) [Planococcus species of South Korea]
- MarottPa1997: pp.114 ( Adult (F) ) [Italian mealybugs on Geranium sp.]
- Koszta1996: pp.169 ( Adult (F) ) [Northeastern North USA]
- WilliaGr1992: pp.393 ( Adult (F) ) [Central and South America]
- Cox1989: pp.9 ( Adult (F) ) [World]
- WilliaWa1988a: pp.163 ( Adult (F) ) [Tropical South Pacific Region]
- Cox1987: pp.74 ( Adult (F) ) [New Zealand]
- CoxBe1986: pp.482 ( Adult (F) ) [Mediterranean basin]
- Willia1985: pp.274 ( Adult (F) ) [Australia]
- McKenz1967: pp.280 ( Adult (F) ) [U.S.A., California]
- Beards1966: pp.432 ( Adult (F) ) [Federated States of Micronesia]
- Beards1960: pp.214 ( Adult (M) ) [Hawaii]
- Ferris1950b: pp.164 ( Adult (F) ) [North America ]
- Borchs1949: pp.95 ( Adult (F) ) [Palaearctic region]
- Zimmer1948: pp.176 ( Adult (F) ) [Hawaii]
- Lawson1917: pp.177 ( Adult (F) ) [U.S.A.: Kansas]
Remarks
- Systematics: GeneBank Accession No. AB439517 (Yokogawa & Yahara, 2009). Tranfaglia (1978), Tranfaglia & Tremblay (1982) and Tremblay et al.(1983) pointed out and evaluated the great morphological affinity (of the adult female) of this species with P. ficus. Rotundo (1975), Rotundo & Tremblay (1975a), Rotundo & Tremblay (1976a), Tremblay et al., (1977) and Rotundo & Tremblay (1980) evaluated its degree of affinity to three mealybug species using female and male morphology, chromosome numbers, endosymbionts, serological methods and immunoelectrophoresis. Danzig & Gavrilov-Zimin (2015) considered Planococcus citri and P. ficus to be synonyms based on variation in morphological characters under different environmental conditions. However, we do not accepted this synonymy since there is strong evidence that they have distinct female sex pheromones (Rotundo & Tremblay 1975) and are closely related but distinct using a combination of morphological and molecular characters (Hardy et al., 2008) Kol-Maimon, et al., 2014, reported that P. citri and P. ficus are easily hybridized in the laboratory, and the hybrids are fertile to some extent, therefore, the possible occurrence of natural populations including hybrids should be taken into account. It is interesting to note that laboratory generated hybrids that resulted from crossing P. ficus females with P. citri males resulted in high mortality, as compared with lower mortality of the reciprocal crossing. Offspring produced as a result of cross-mating between P. ficus females and P. citri males hardly ever survive in the laboratory, therefore, P. ficus males displaying characteristics of P. citri males are unlikely to complete their development on plants in the field. In addition to the five haplotypes detected in Turkey, an additional nine haplotypes were detected worldwide, giving 14 haplotypes in total. With respect to geographic distribution, there were three haplotypes from the Americas, six from the Middle East, five from Europe, four from the Far East and three from Africa. Hap 1 and 2 are the two main groups according to haplotype network and both haplotypes are distributed all over the geographic groups except Hap 2, which was not detected in South Africa. The study indicates that Hap 1 and 2 are possibility different cryptic species based on few base changes, because of different host plant range. Different cryptic species might not be so significant for chemical control tactics. However, they might be important for biological control strategies, because of their effect on fitness cost of parasitoids. (Karagaoğlu, et al., 2020) Conflicting data was found in public databases when two sequences of Planococcus minor and two of "Planococcus citri" shared the same haplotype (Oliviera et al., 2023). Caballero & Kondo (2024) considered Planococcus citri-minor a species complex.
- Structure: Colour photograph by McKenzie (1967), Schmutterer (1990), Carvalho & Aguiar (1997), Wong et al. (1999) and by Matile-Ferrero et al.(2004). Several aspects of the male chromosome system studied by Nur (1966, 1967, 1968). SEM micrograph of trilocular pore and of wax secretions given by Foldi (1991). Ultrastructure of the symbionts studied by Iaccarino & Tremblay (1971). Rotundo and Tremblay (1974) described a technique for extractions of the female sex pheromone, and Rotundo & Tremblay (1982a) gave preliminary data about its activity. Rotundo & Tremblay (1975b) evaluated the attractiveness of the female sex pheromone. The female sex pheromone was isolated, identified and synthesized by Bierl-Leonhardt et al. (1981). Female sex pheromone synthesized by Gaoni (1982), Carlsen & Odden (1984) and by Wolk et al. (1986). Rotundo & Tremblay (1980a) evaluated the daily rate of release of the female sex pheromone.
- Biology: James (1937a) showed that reproduction in sexual (in laboratory population in England) and evaluated its sex ratio. James (1938) proved that environmental humidity has no decisive effect (in laboratory population in England) on sex ratio. Nelson-Rees (1960) studied the life history and sex predetermination. Nelson-Rees (1961) described the modification of the ovary due to aging. Rotundo & Tremblay (1975c) reported on the attractiveness of virgin females to hymenopterous parasites. Rotundo & Tremblay (1982b) reported on hybridization between this species and Pl. ficus. Tranfaglia (1978), Tranfaglia & Tremblay (1982) and Tremblay et al. (1983) pointed out and evaluated the great morphological affinity (of the adult female) of this species with P. citri. Rotundo & Tremblay (1982a) bioassayed the synthetic female sex pheromone. The female sex pheromone and some analogues were field-bioassayed by Dunkelblum et al. (1987). Moreno et al. (1984) studied the complex hierarchy of male responses elicited by the female sex pheromone. Ortu & Delrio (1982) presented observations on the use of the pheromone in control of this species. Gothilf & Beck (1966) developed an artifical, defined diet for laboratory rearing of this mealybug. Campbell (1983) assessed the presence and spatial distribution on cocoa trees in Ghana. Natural enemies discussed by Bartlett (1978) and by Moore (1988). Parasites in Palaearctic region discussed by Trjapitzin (1989). Smith et al. (1997) reported from citrus in Australia, at least 6 annual generation (in Queensland and the Northern Territory), 4-5 gnerations in New South wales, while 3-4 in Victoria and South Australia. Mortality of Planococcus citri (Risso), was higher on green than on red or yellow variegated Coleus blumei ‘Bellevue’ (Bentham) plants, and developed faster with a higher fecundity on red variegated plants. (Qin, et al., 2011) Mean longevity for adult males is 73.3 ± 1.6 h. Planococcus citri (Risso) males may fertilize an average of nine females, with a maximum of 27 females during male life span. There is a sexual maturation period of the adult males which lasted for about 42 h., indicated by maximum growth of waxy caudal filaments. (Mendel, et al., 2012) The highly unusual reproductive system of mealybugs has made them a popular model for studying unusual chromosome behavior, with a rich history of cytogenetic research. They are a promising system for studying genomic imprinting (parent-of-origin specific gene expression) and non-Mendelian inheritance. P. citri has also been a valuable model for understanding organellogenesis, the evolutionary process by which endosymbiotic bacteria transition into cellular organelles such as mitochondrial and chloroplasts. (Ross, et al. 2024)
- Economic Importance: This is one the most cosmopolitan mealybugs, damaging many outdoor crops in the tropics and subtropics, as well as in greenhouse in the temperate regions (Bodenheimer, 1930a, 1951a; McKenzie, 1967; Bartlett, 1978e; Cox, 1989). A sporadic olive pest in the Mediterranean region (Swirski, 1985). Rosciglione & Castellano (1985) showed that this mealybug transmitted Grapevine Virus A (GVA) from grapevine to Nicotiana clevelandii.
Lagowska (1995) reviewed the prospects for biological control in greenhouses.
Hanna et al., (1956) reported that P. citri is a vector of the swollen-shoot virus disease of cacao in Ghana.
Cabaleiro & Segura (1997) studied in Spain the field transmission of Grapevine Leafroll Associated Virus 3 (GLRaV-3) by the mealybug Planococcus citri; it is very likely that the studied mealybug was actually Planococcus ficus (Signoret).
The provisioning of alternative sugar sources with sugar-feeders has been shown to disrupt the mutualistic association between ants and honeydew producers. (Pérez-Rodríguez, et al., 2021)
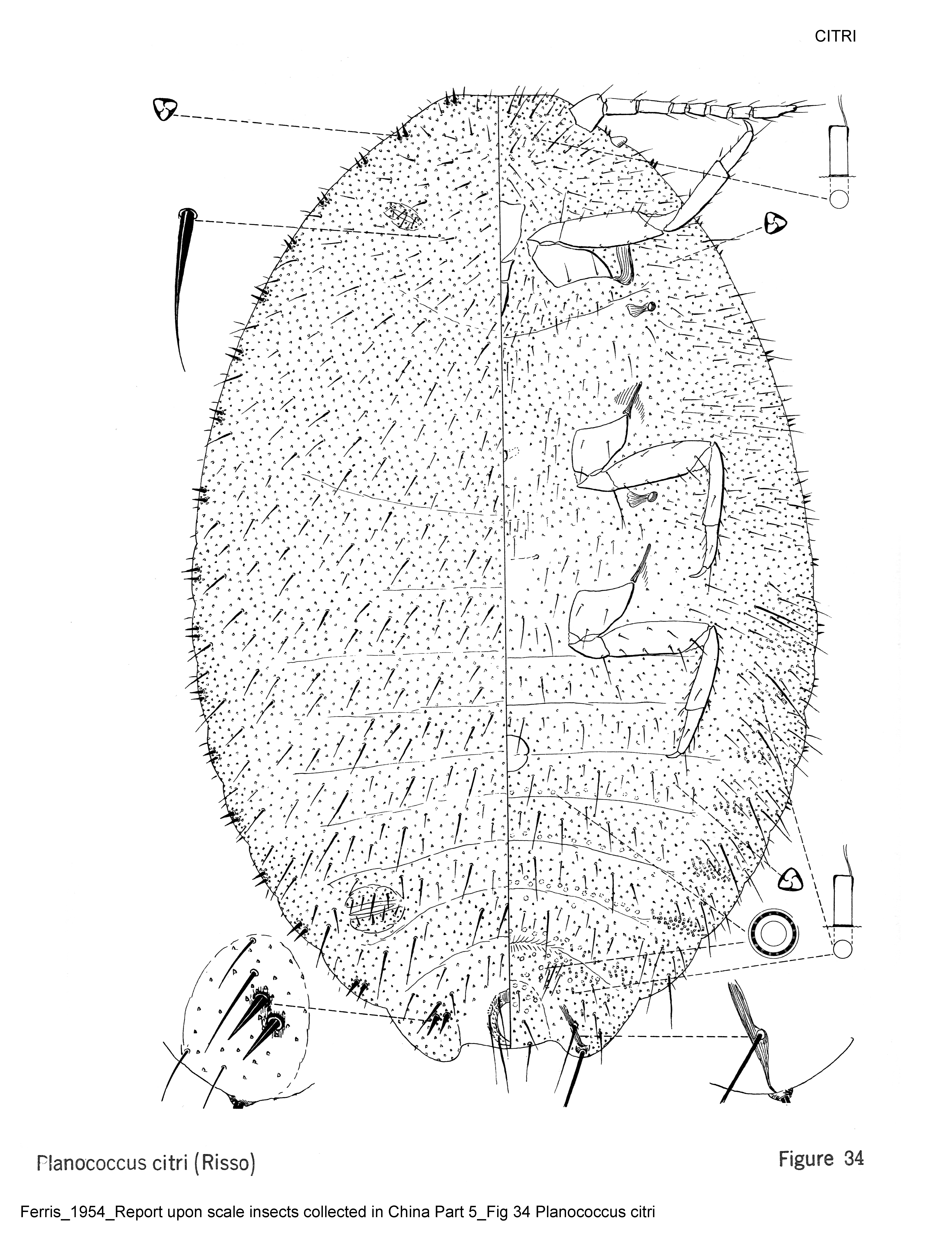
- General Remarks: Description and illustration of adult female by Ferris in Zimmerman (1948), Ferris (1950b), McKenzie (1967), Cox (1981; 1987; 1989), Williams & Watson (1988a), Williams & Granara de Willink (1992), Williams (2004a) and by Danzig & Gavrilov (2010). Good description and illustration of the adult male given by Beardsley (1960) and by Afifi (1968). Description of adult female by Borchsenius (1949). Cox (1981) evaluated the effect of environmental conditions on the range of variation of certain morphological characters in the adult female.
Illustrations




Citations
- AbdElKDaKo1988: biological control, chemical control, 270-275
- AbdRab2001d: biological control, distribution, 1362
- AbdRab2012: list, 613
- AbdRabEv2021: distribution, host, 43
- AbdelkAfSm1998: biological control, distribution, economic importance, host, life history, 483-488
- AbdulRHe2018: distribution, host,
- AblesRi1981: biological control, economic importance, 273
- AbrokwDzGa2016: vector, virus,
- AbuAllOmRa2025: natural enemies, 42
- AcevedLoGo2025: distribution, host, 141
- Ackono2002: biological control, 105-112
- Afifi1968: description, illustration, taxonomy, 93-98
- AhlawaPa2003: disease transmission, 447-474
- AhmedAb2010: biology, distribution, host, natural enemies,
- AhmedApKa2023: control, natural enemies, virus transmission, 743
- AhmedPoMi2025: distribution, key, list of species, morphology, 3, 9
- AlAnsiAlFa2020: natural enemies, 70, 71
- AlJbouHe2025: biological control, natural enemies, 878
- AlboreLoGr2024: chemical ecology, control, host,
- Aldric1996: chemical ecology, chemistry, life history, physiology, 205-206
- Alfons1875: description, distribution, host, taxonomy, 428-431
- AllouiChAt2022: control,
- AllwinRaRa2025: biological control, ecology, economic importance, host, illustration, natural enemies,
- Almeid1973b: distribution, host, 19
- AlmeidDaBe2013: Grapevine leafroll disease transmission, 3
- AlmeidPeMa2018: distribution, host, 356
- AltierNi1999: biological control, 975-991
- AlvareLiLu2020: phylogeny, 5
- AndradBoPe2017: biology, disease transmission, economic importance,
- AngeleHeCe1992: biological control, distribution, host, 27-29
- Anneck1969: biological control, 849-854
- AnneckPr1976a: biological control, distribution, host, 105-108
- Arai1996: distribution, ecology, host, life history, 25-34
- Arai1997: distribution, ecology, host, life history, 263-266
- Archan1930: distribution, host, 78
- Argyri1983: distribution, host, 365
- ArgyriStMo1976: biological control, distribution, host, 22
- ArmandKhNa2024: entomopathogenic fungi,
- Armita1919: biological control, 257-260
- ArouaKaBe2019: distribution, host,
- Ashmea1879: description, distribution, host, taxonomy, 160
- AssefaMa2018: DNA sequencing, distribution, host,
- Astrid2001: economic importance, host, 76
- AttiaMaAb2022: control, natural enemies,
- AtwaMoSh1987: chemistry, life history, taxonomy, 1655-1676
- AvasthSh1987: description, distribution, host, illustration, taxonomy, 36-38
- Avidov1961: chemical control, distribution, host, life history, 144-148
- AvidovErGo1960: chemical control, economic importance, 33-45
- AwadalBaEl2023: biological control, natural enemies,
- Ayouta1940: distribution, host, 2-4
- Ayouta1940: host, 2-4
- Bain2016: genetics, reproduction, 16
- BainMaFi2021: chemistry, chromosome, genetics,
- BakryWaHe2025: mechanical control, 15
- Balach1927: distribution, host, 187
- Balach1928a: taxonomy, 143
- Balach1932b: ecology, 517-522
- BalakrViGo1987: biological control, 59-61
- BalalaShSh2025: biological control, chemical control, natural enemies,
- BalciuBuPu1995: biological control, 247-267
- BalikaKoPr2011: distribution, host, 580
- BansalFe2025: control, genetics, resistance, 8
- BarberPoBo2025: biological control, entomopathogenic fungi, 2, 8
- Barnes1935: biological control, distribution, host, 526-527
- Bartle1957: biological control, 753-755
- Bartle1969: biological control, 875-878
- Bartle1974: biological control, 553-556
- Bartle1978e: biological control, distribution, economic importance, host, 150-155
- BartleLl1958: distribution, economic importance, host, 90-93
- BasheeAsRa2014: biological control, distribution, host, 49, 51
- Basing1931a: biological control, distribution, economic importance, host, 6-8,30-31
- BaskarLaUt1999: biological control, 605-606
- BatsanKaKi2017: distribution, host, 265
- BattagTrFr1997: biological control, 3-11
- Beards1960: description, distribution, host, illustration, taxonomy, 224
- Beards1966: distribution, host, taxonomy, 432-434
- Beatty1944: distribution, host, 114-172
- Beccar1971: distribution, host, 195
- Beders1969: chemical control, distribution, host, 933-940
- Beingo1957: biological control, distribution, economic importance, host, 23-25
- BeltraAdAv2015: distribution, host, 5
- BeltraGaSo2013: 1492
- BeltraLlHu2025: distribution, economic importance, host, key, survey, 108
- BeltraSoMa2012: molecular data, phylogenetics, 167-171
- BenDov1994: catalog, 355-358
- BenDov2006a: distribution, host, 206
- BenDov2012: catalog, distribution, host, 36, 44
- BennetRoCo1976: biological control, economic importance, 359-365
- BennetSi1964: biological control, 81-94
- BernarVi1999: biological control, chemical control, 81-84
- BertelBa1966: distribution, host, 17-46
- BesterBrGi2026: diagnosis, economic importance, genome, host, phylogeny,
- Betrem1937: description, distribution, host, illustration, taxonomy, 64-69
- BierlLMoSc1981: chemistry, physiology, 389-392
- Bishop1931: biological control, distribution, host, 1-7
- BlachiDr2000: chemical control, distribution, host, 316-318
- Blumbe1997: biological control, ecology, 225-236
- BlumbeFrSu2002: biological control, life history, structure, 385-395
- BlumbeGrSt1997: biological control,
- BlumbeKlMe1995: biological control, life history, structure, 157-163
- BlumbeVa2001: biological control, life history, 191-199
- BluthgFi2002: ant association, 796,797,799,
- Bodenh1924: description, distribution, host, taxonomy, 78-81
- Bodenh1926: distribution, host, 45
- Bodenh1928a: biological control, distribution, economic importance, host, 6-14
- Bodenh1930a: economic importance,
- Bodenh1937: distribution, host, 219
- Bodenh1943: distribution, host, 14
- Bodenh1951a: biological control, chemical control, description, ecology, economic importance, illustration, life history, physiology, structure, taxonomy, 396-454
- Bodenh1953a: distribution, host, 119
- Boisdu1867: taxonomy, 348
- Bolu2012: biological control, 110
- BongioMaMa2001: life history, structure, 3809-3817
- Borchs1937a: description, distribution, host, illustration, taxonomy, 162-164
- Borchs1949: description, distribution, host, life history, taxonomy, 131-134
- Borden1923: biological control, distribution, host, life history, 1-10
- Bouche1844: description, distribution, host, taxonomy, 301-302
- BouhelDeDe1932: control, distribution, host, 1-60
- Boyce1948: control, distribution, economic importance, host,
- Brader1979: biological control, chemical control, 225-254
- BrahmaKoGu2018: chromosome, cytogenetics, endosymbiont, genetics, life cycle,
- BrahmaKoGu2018: chromosome, genetics, molecular biology,
- BrainKe1917: distribution, 182
- BrancoSoBe2008: biological control, life history, 273-274
- Branig1916: biological control, 304-306
- Brimbl1962: distribution, economic importance, host, 227-228
- Brock1925: economic importance, 349,366
- BroshToNe1989: chemical control, 1464-1465
- BruntKe1971: disease transmission, economic importance, host, 591-602
- BuangAmAd2025: host, virus, virus transmission, 11-12
- Buchne1953: structure, taxonomy, 230-238
- Buchne1965: structure, taxonomy, 255-261
- BugajNJuKa2021: distribution, host, list, 310
- BugilaFrSi2014: behavior, biological control, evolution, 439–453
- BugliaFe2004: cytology, genetics, 284-294
- BugliaPrFe1999: chemistry, life history, structure, 71-73
- CABI1969: distribution,
- CABI1999: distribution,
- CabaleSe1997: ecology, economic importance, 283-287
- CaballKo2024: host, illustration, key, morphology, taxonomy, 2, 9, 20
- CaballKoRa2023: distribution, host, survey, 36
- CaballRaGi2018: distribution, host, 120
- CalabuGaPe2014: ant association, 406
- CalisiKlKa2005: natural enemies,
- Calkin1983: distribution, economic importance, 321
- Campbe1983: distribution, ecology, host, life history, 137-151
- Campbe1984: distribution, ecology, host, life history, 163-174
- CamposMaRi2003: biological control, life history, 139-147
- CamposMaRi2003: biological control, 139-147
- CanhilUyCa2001: biological control, 117-125
- Capra1927: biological control, distribution, host, 152-160
- CarlseOd1984: structure, 501-504
- CarmoSiTo2026: natural enemies, 3
- CarmonBeMe2025: ecology, life cycle, male, 2, 9
- CarrasCaTo2025: phylogeny, virus, 3, 5
- Carrer1979: biological control, distribution, host, 521-526
- CarrilHoAl2024: distribution, economic importance, host, 549, 562
- CarvalAg1997: description, distribution, economic importance, host, life history, 168-170
- CarvalFrAg1996: distribution, economic importance, host, 614
- CaustoPeSi2006: distribution, 138
- CavaliMaTr2008: molecular data, taxonomy, 65-68
- CeballWa2004: biological control, host, 235-244
- CebeciAr2006: distribution, host, 141
- ChackoKrAn1978: biological control, 14-19
- ChackoKrRa1976: chemical control, 118-119
- ChamLuRo2019: distribution, host, 361
- ChellaRaBi2026: biological control, natural enemies, 39
- ChenShCh2024: molecular biology, 3
- ChibirRuTk1991: chemical ecology, chemistry, 1209-1213
- ChirinAnCa2021: natural enemies, 13
- ChoiLe2022: endosymbiont, phylogeny, 358,359
- ChongOe2006a: biological control,
- ChongOe2007a: biological control, host distribution, 933-944
- ChongOeOs2005: biological control, distribution, host, life history, 39-48
- Chou1947a: chemical control, 38
- ChowdhRa2024: distribution, economic importance, host, phenology, survey, 6, 10, 12
- CidFe2010: Grapevine leafroll disease transmission,
- CidPeCa2010: disease transmission, distribution, economic importance, host, 619-630
- Clause1958: biological control, economic importance, 291-310
- Clause1958a: biological control, distribution, host, 443-447
- Cloyd1999: biological control, ecology, life history, 1-60
- Cloyd2003: chemical control, 1397-1399
- CloydHe2023: biological control, entomopathogenic fungi,
- Cocker1893n: distribution, host, taxonomy, 267
- Cocker1894z: distribution, 210
- Cocker1896b: taxonomy, 325
- Cocker1897: distribution, host, 200
- Cocker1898g: taxonomy, 109
- Cocker1899n: distribution, host, 7
- Cocker1902p: taxonomy, 252
- Cocker1902z: distribution, host, 614-615
- CoffeeBoRe1986: distribution, host, 1-73
- Cole1933: biological control, distribution, host, 855-864
- Comper1938: biological control, 315-337
- Comper1939: biological control, 57-73
- Comper1939d: biological control, distribution, host, 13-15,
- Comper1969: biological control, 755-764
- ComperZi1955: biological control, 94-116
- Comsto1881a: description, distribution, host, illustration, taxonomy, 342-343
- ContreMoMa2021: natural enemies,
- Cook1915: control,
- CookHo1908: catalog, diagnosis, distribution, host, 30
- CoplanPeHe1992: biological control, distribution, host, 44-47
- CorreaLoMa2015: distribution, genetic structure, host, molecular biology,
- CoryHi1959: chemical control, 344-345
- CotoSa2001: distribution, host, 60-68
- Cox1981: taxonomy, 47-53
- Cox1983: illustration, morphology, taxonomy,
- Cox1987: description, distribution, host, illustration, taxonomy, 74, 193
- Cox1989: description, distribution, host, illustration, taxonomy, 15-17
- CoxBe1986: taxonomy, 482-483
- Crouze1973: biological control, distribution, host, 15-39
- CulikMaVe2007: distribution, host, 1-5
- DaSilvMoBa2015: biology, life cycle,
- DahlstHa1999: biological control, 919-933
- DanzigGa2010: description, distribution, host, illustration, taxonomy, 42-43
- DanzigGa2015: description, distribution, host, illustration, key, taxonomy, 207-211
- DanzigKo1990: distribution, host, 38-52
- DarvasSz1987: chemical control, distribution, host, 343-351
- DaviesCeWa2004: biological control, 181-188
- Davis1908: diagnosis, reproduction,
- DeBachDiFl1951: biological control, 347-348
- DeBachDiFl1951: biological control, 1,14
- DeBachHu1971: biological control, 113-140
- DeBachHuMa1976: biological control, 255-285
- DeFlui1941: distribution, economic importance, host, life history, 1-119
- DeLott1967a: distribution, host, 119
- DelGue1906: distribution, host, 257-263
- DelaCrCh1996: biological control, distribution, economic importance, host, 100-104
- DelabiCa1991: distribution, economic importance, host, 53-57
- DelabiDaCa1994: ecology, economic importance, life history, 91-103
- DelrioOrUs1979: biological control, distribution, host, 539-541
- DemardBaDi2025: behavior, histology, host, 2, 5, 12-13
- DemontOrCo2007: molecular data, taxonomy, 59-64
- DenninDu1954: chemical control, 310-316
- DewerAbSc2018: DNA, distribution, host, morphology, 155-159
- DhaunrWaJi2019: biological control, economic importance, 1
- DineshVe2016: biological control, natural enemies,
- Domeni1951: biological control, distribution, host, 157-180
- Doutt1949: structure, taxonomy, 835
- Doutt1951: biological control, chemical control, distribution, economic importance, host, 37-40
- Doutt1952: structure, taxonomy, 391-397
- Doutt1952a: biological control, distribution, economic importance, host, 343-344
- DowellGiJe2016: distribution, 119
- DownieGu2004: distribution, host, molecular data, phylogeny, 258-259
- Dozier1926a: biological control, 97
- DreistClFl1994: control, description, economic importance, life history, taxonomy,
- DuWaLi2025: evolution, genome, phylogenetics, taxonomy, 4
- DuncanMa2024: biological control, economic importance, host, life cycle, phenology, 277, 283
- Dunkel1999: chemical ecology, chemistry, 251-276
- DunkelBeGo1987: chemistry, structure, 863-871
- DunkelBeGo1987a: chemistry, structure, 149-154
- DunkelZaGr2002: chemical ecology, chemistry, 213-220
- Dziedz1990: distribution, host, 34-35
- EPPO2024: control,
- EPPO2024a: control,
- ESKSAE1994: distribution, list, 111
- EhsasAyIk2024: genome, molecular biology, 68
- ElAalaRaKa2024: biological control, 2
- ElAalaRaSb2024a: biological control, entomopathogenic fungi, 1081
- ElAalaSb2025: natural enemies, 14
- ElangoSr2017: ecology, illustration, 379
- ElekciKa2021: distribution, host, 10
- ElekciSe2007: biological control, distribution, host, 29-34
- ElimemJaBo2023: control,
- ElimemJeLa2022: distribution, host, illustration, natural enemies,
- ElzenKi1999: biological control, 253-270
- Epstei1991: chemistry, chromosome, structure,
- ErensoBlGo2023: phylogeny, 4
- ErkiliDe2008: biological control, distribution, host, 297-298
- ErlerErTo2008: biological control, chemical control, 297
- Essig1909a: distribution, taxonomy, 46
- Essig1910b: distribution, host, 339-345
- Essig1910f: biological control, 260-274
- Essig1910g: chemical control,
- Essig1911b: natural enemies, 390
- Essig1915a: biological control, description, distribution, host, life history, taxonomy, 126-129
- EtzelLe1999: biological control, 125-197
- EvansAb2023: distribution, host, illustration, key, taxonomy, 6, 7, 9, 23, 66
- EwartMe1956: chemistry, 441-447
- Ezzat1987: structure, 57-62
- EzzatMc1956: description, distribution, host, illustration, taxonomy, 55, 65-67, 71
- EzzatNa1987: distribution, 89
- FallahJa2017: distribution, host, natural enemies, 6, 10, 34
- FargerMo1974: biological control, chemistry, life history, 26-28
- Fernal1903b: 99-100
- Fernan1992: distribution, host, 61
- FernanMiPo2025: ecology, honeydew, natural enemies,
- FerrarBuPr1998: chemistry, structure, 45
- FerrarBuRo2001: structure, 93-101
- Ferris1950b: description, distribution, host, illustration, taxonomy, 165-167
- Ferris1953a: distribution, host, taxonomy, 494
- FieldLyMa2004: molecular data, structure, 109-115
- FilhoGrBo2008: biology, life history,
- FiliaFeRo2019: genetics, reproduction,
- FiliaRo2016: evolution, genetics, reproduction,
- Fisher1963: biological control, 1-39
- FlaherWi1999: biological control, 853-869
- Flande1937: biological control, 401-422
- Flande1943: biological control, 117,128,137
- Flande1949a: biological control, 257-274
- Flande1951b: biological control, 93-98
- Flande1953a: biological control, distribution, economic importance, host, 266-269
- Flande1958a: biological control, 579-584
- Flande1959b: biological control, 125-142
- Foldi1991: structure, 163-188
- Foldi1997: phylogeny, taxonomy, 205
- Foldi2000: distribution, host, 78
- Foldi2001: distribution, 303-308
- Foldi2003: distribution, host, 149
- FoldiGe2018: distribution, list, 13
- FoldiKo2006: distribution, host, 308
- FoldiKo2006: distribution, host, 312
- FranciKaRo2012: host, 295
- Franco1996: control, ecology, life history, 711
- FrancoAnLe2008: chemical ecology, ecology, life history, 230-231
- FrancoCoLu2021: reproduction, 254
- FrancoMa2001: distribution, host, 191-196
- FrancoPa1991: distribution, host, 282-285
- FrancoRuMa2011: distribution, 20,25
- FrancoRuSu2002: chemical ecology, ecology, economic importance, life history, 297-303
- FrancoRuSu2005: biological control, life history, 232-233
- FrancoRuSu2005: chemical ecology, ecology, life history, 232-233
- FrancoSiBr2016: biological control, natural enemies, 72
- FrancoSiCa2000: biological control, distribution, economic importance, host, illustration, life history, taxonomy, 1-142
- FrancoSiCo2008: chemical ecology, life history, 122-130
- FrancoSiFo2011: biological control, chemical ecology, natural enemies,
- FrancoSuBo2004: biological control, 507-522
- FrohliRo1970: distribution, economic importance, host, 1-10
- Fursch1967: biological control, 309-310
- FustLaSh2025: GLRV, vector, 2988
- GaimarGoEl2024: host, natural enemies, 561
- GallegUrRu2026: control, molecular biology, 2, 8
- Gaoni1982: structure, 5215-5218
- Garcia1998JF: biological control, host, life history, 1-86
- GavaraVaNa2024: control, 8
- GavaraZaMa2025: behavior, biology, chemical control, male, 7
- GeorgaBuHo1972: chemical control, 16-22
- Germai2011: distribution, economic importance, 31-34
- Germai2011a: distribution, economic importance, 8
- GermaiDeLa2016: host, list of species, 4
- GermaiMaPi2002: distribution, host, 255
- GermaiMiPa2014: distribution, 24
- Gertso2016: distribution, host, 153
- GertssSc2020: distribution, host, 137
- GharizHe2003: biological control, distribution, host, 85-86
- GhoshMe2024: diagnosis, honeydew, host, host distribution, illustration,
- GodoiSoMe2024: host, record,
- Golber1982: biological control, 86-90
- GollifJaDa1977: distribution, host, 171-177
- GomesCRe1949: distribution, host, 63-64
- GomezC1932: biological control, 1-59
- GomezC1950: biological control, distribution, host, 1-18
- GomezC1951: biological control, distribution, host, 21-28
- GomezC1954a: biological control, distribution, host, 19-35
- GomezM1929: taxonomy, 5
- GomezM1937: description, distribution, host, taxonomy, 333-339
- GomezM1958c: distribution, host, 407
- GontijToAb2025: biological control, ecology, history, natural enemies, 3
- Gonzal1969: biological control, 839-847
- Gonzal2011: chemical control, description, distribution, illustration, life history, taxonomy, 135-143
- Gonzal2016: description of male, diagnosis, distribution, host, illustration, taxonomy, 14, 240, 247, 296-305
- GonzalVo2004: distribution, economic importance, host, 41-62
- GoszczGo2011: biological control, distribution, host, 109-110
- GothilBe1966: behavior, life history, physiology, 489-490
- Goux1931: distribution, host, 4
- GovindJoSr2003: biological control, 96-102
- GracyReNe2025: virus transmission, 7
- GradyRe1940: distribution, economic importance, host, taxonomy, 1-32
- Granar1979: description, distribution, host, illustration, taxonomy, 143-149
- Granar1991a: distribution, host, 267-268
- GranarCl2003: distribution, host, 625-637
- GranarScTe1997: distribution, economic importance, host, 97-98
- Graven2003: biological control, 71-82
- Gray1954: life history, 174-176
- Greath1971: biological control, distribution, host,
- Greath1976: biological control, economic importance,
- Green1930c: distribution, host, 280
- GriffiTh1947: chemical control, distribution, host, 386-388
- GrossBiGa2001: biological control, chemical control, 219-221
- GrossDrBl1999: distribution, economic importance, host, 463-468
- GrossDrSt1997: biological control, distribution, economic importance, host,
- GrossDuAs2002: chemical ecology, economic importance, life history, 283-295
- GroveDe2017: distribution, host, 300
- GroveDeDa2013: distribution, host, 378
- GugliuMaMa2019: distribution, host, list, 4, 7
- GuindaNoWo2017: ant association,
- Gullan2000: description, taxonomy, 160-166
- GullanDoKn1987: distribution, host, taxonomy, 52
- GuptaKoGa2021: bacteria, control,
- GutBr1994: chemical ecology, control, 67-75
- GyamweDoAk2023: virus transmission, 12
- HagenBoMc1976: biological control, life history, 93
- HakkonPi1984: biological control, 1109-1121
- HalimaGeMd2014: distribution, host, 1-2
- HalimaZoCh2025: distribution, host, illustration, survey, 78-79
- Hall1926a: distribution, host, 6
- Hall1927d: distribution, host, 267-287
- Hall1937: distribution, host, 126
- Hall1969: economic importance, 823-826
- HallFo1933: distribution, economic importance, host, 1-55
- HamidMi1996: biological control, distribution, host, 711
- HannaHeJu1952: chemical control, 334-335
- HannaJuHe1955: chemical control, distribution, economic importance, host, 889-710
- HannaJuHe1956: control, disease transmission, economic importance, 219-227
- HarbiAbBr2025: distribution, economic importance, 2
- HardyGuHo2008: molecular data, phylogeny, taxonomy, 51-71
- HarishAr2022: endosymbionts, 244
- HefetzTa1990: chemical ecology, chemistry, life history, 502-506
- HegdeChYa2024: distribution, host, 472
- HeidarJa2000: biological control, 49-53
- HelalDoEl2000: distribution, host,
- Hellri2004: distribution, host, 83
- Hempel1900: description, distribution, host, illustration, taxonomy, 389-390
- Hempel1901: distribution, host, taxonomy, 110-111
- HennekOpAl1987: biological control, distribution, host,
- HerbetRo2025: chromosome, genome, male, reproduction,
- HernanNiMa2011: host, 379-380
- HerrbaAlPr2017: Grapevine leafroll disease transmission, 486
- HerricCl2023: chemical control,
- HeyaKhKe2020: genetics,
- Hodgso1969c: biological control, distribution, economic importance, host, life history, 22
- Hodgso2012: taxonomy, 64
- Hodgso2020: key, 34
- HodgsoHi1990: distribution, host, 2-22
- HodgsoHi1991: distribution, host, 144
- HoffmaRiSh1998: biological control, 268-293
- HogendClSw2006: host, life history, physiology, 201-211
- Hollin1923: description, taxonomy, 52, 53
- HosseiHa2011: biological control, 46
- HoyHe1985: biological control,
- HuHeWa1992: description, distribution, host, taxonomy, 179
- HuangLvDi2025: evolution, genome, phylogeny,
- HusseyBr1971: biological control, distribution, host, 95
- IaccarTr1971: structure, 35-48
- IbrahiGuWa2021: chemical control, chemistry, symbiont,
- Iperti1961: economic importance, 14-30
- IrenAtOk1973: chemical control, 1
- IsaqovZoHu2024: behavior, diagnosis, distribution, history, host, 27, 31
- IslamJa1992: biological control, 17-22
- IslamJa1993a: biological control, life history, 322-324
- IzraelLaNe2020: Wolbachia, natural enemies,
- IzraelWoMo2025: biological control, natural enemies, resistance, virus, 2
- JacasMGa2001: biological control, chemical control, 103-112
- Jackso1995: biological control, life history,
- JacobsReCr1978: chemistry, 448-457
- JalaliSiBi2000: biological control, life history, 159-162
- JalilvShFa2014: distribution, host, natural enemies, 3, 4
- James1937a: life history, 429-461
- James1938: life history, 73-79
- Janezi1954: distribution, host, 127
- JansenAl2023: dispersal, host, 30,35
- JaposhYeKa2023: natural enemies, 577=582
- JaszaiDa1983: chemical control, 198-202
- JayaraChGo1988: biological control, 77-78
- Jeppso1969: chemical control, economic importance, physiology, 917-921
- JiSu2012: distribution, host, 1-5
- JohnsoGi2010: distribution, economic importance, host, life history, 354-359
- JoshiSuRe2021: diagnosis, distribution, host, illustration, key,
- JoyceHoBe2001: biological control, life history, 49-57
- Kaneko2025: distribution, ecology, host, natural enemies, phenology, 1
- KansuUy1979: biological control, distribution, host, 565-567
- Kaplan1996: biological control, 215-216
- Kaplan1996: biological control, chemical control, 215-216
- Kaplan1998: biological control, chemical control, 165-176
- KaracaKaSi2016: biological control, economic importance, entomopathogenic fungi, host, illustration,
- KaracaSa2017: biology, heat-treatment, reproduction,
- KaracaSaSm2020: DNA, dispersal, host, phylogenetics,
- Katsoy1996: biological control, chemical control, economic importance, life history, 21, 83-85
- Kaussa1957: distribution, host, 2
- KavyaYHeBh2026: ant association, host, 391-392
- Kawai1972: distribution, host, 8
- Kawai1987: distribution, host, 77
- KawaiMaUm1971: distribution, host, 15
- KawpetGa2024: biological control, distribution, economic importance, host, illustration, natural enemies, survey,
- KaydanJa2010: host, natural enemies, 468
- KaydanKiKo2005: distribution, host, 91
- KaydanUlEr2007: catalog, distribution, host, 102
- KetterVeBi2003: biological control, distribution, host, 199-206
- KhaladBoMa2024: distribution, species richness, survey,
- KhalafZhAd2023: virus transmission,
- KhanSaAk2012: biological control, 3, 6
- KhattaMu2026: biological control, chemical ecology, host, natural enemies,
- KimPaLe2021: chemical control,
- King1900b: taxonomy, 214-215
- KingLe1984: biological control, 1
- KingMo1984: biological control, 206-222
- KinjoNaHi1996: distribution, host, 126
- Kiritc1932a: taxonomy, 263-265
- Kiritc1936: distribution, host, 73, 74
- KoffiBaDe2025: DNA, ant association, distribution, host, natural enemies, virus transmission, 11
- KohliGuNa2021: genetic structure, symbionts,
- KolMaiGhFr2014: DNA, genetics, reproduction,
- KolMaiMeFr2014: male, reproduction,
- Kondo2001: distribution, host, taxonomy, 37
- Kondo2008a: distribution, host, 25-29
- KondoRaVe2008: distribution, host, 40-41
- KondoWa2022a: distribution, host, list, 24
- KondoWa2022d: illustration, 549
- KontodLyKa2005: biological control, life history, 359
- KosztaKo1988F: distribution, host, 60
- Koteja1974b: distribution, taxonomy, 74
- KozarFoZa1996: distribution, host, 64
- KozarSzFe2013: distribution, list of species, 92, 107, 125, 144, 161
- KreiteAuGe2006: distribution, economic importance, host, 143
- Kriegl1975: chemical control, distribution, host, 71-77
- Krishn1988: biological control, 97-98
- KrishnMa1989a: biological control, economic importance, 466
- KrishnMa1994: distribution, economic importance, host, 12-13
- KumarMaSe2016: distribution, host, 643
- KumaraKoRa2024: biological control, natural enemies, 3
- KutukKaTu2014: biological control, chemical control, distribution, economic importance,
- Kuwana1907: distribution, host, 184
- KwonDaPa2003a: distribution, host, taxonomy, 406-408
- KwonHaCh2005: distribution, host, 54-60
- LaChanScBu1967: biological control, chemical control, life history, 147
- LaflinGuPa2004: distribution, host, 475-477
- LaflinPa2004: distribution, host, life history, 982-988
- LafonCoHu1961: economic importance, 37-43
- Lagows1995: biological control, distribution, economic importance, host, 5-10
- Lagows1995a: biological control, distribution, economic importance, host, 375-378
- LagowsGo2020: distribution, 70
- LagowsGoHo2022: distribution, host, list, 11
- Laing1927: distribution, host, 36
- Laing1928: distribution, host, 214-215
- LaneesSuBa2020: biological control, natural enemies, 459
- Lashin1956: distribution, 27-28
- Lawson1917: description, distribution, host, illustration, taxonomy, 177-178
- LeMaguFuCh2013: economic importance, host, 416
- Leach1940: life history, 1-3
- LedieuHeDe1989: economic importance, 405
- LeeSuKi2025: chemical control, physiology,
- LeiZaLe2000: distribution, economic importance, host, 43-44
- LeiteAgCh2025: economic, mechanical control, vector, virus, 1
- LentinMuMu2018: control, heat-treatment, reproduction, 264
- LeonKo2017: biological control, diagnosis, host, illustration, natural enemies, 77-78
- LeviZeFeDa2014: control,
- LiesenMaDe2025: genome, molecular biology, 5, 7
- LincanHoCa2010: host, 6
- Lindin1932f: taxonomy, 201
- Lindin1939: distribution, host, 37
- LiottaMiRa1977: economic importance, host, 29-67
- LiuChCh2020: DNA sequencing,
- LiuChCh2020: DNA sequencing, phylogenetics,
- LiuChCh2020: DNA sequencing, phylogenetics,
- LiuLiWa1995: chemical ecology, chemistry, 3837-3843
- Lloyd1958: biological control, 450-461
- Lloyd1966: life history, physiology, 265
- LongoMaPe1995: distribution, 120
- LopesFiMa2008: distribution, host, 153-154
- LopesOlOl2023: chemical control, control,
- LopezMBeRe2014: endosymbionts,
- Lorick1970: structure, taxonomy, 11-30
- Lorick1970a: structure, taxonomy, 1-68
- LuckFoSc2003: biological control, 179
- MadihRaZa2021: biological control, natural enemies,
- MahfouDh2009: biological control, distribution, host, 154-160
- MalagnPeDu2005: molecular data, taxonomy, 349
- MalausFeWa2011: distribution, molecular data, phylogeny, 142-155
- MalipaDuSm2000: biological control, 79,88,89
- Malump2011a: distribution, host, illustration, 55-57
- Malump2012b: distribution, host, 211,213
- Malump2014a: distribution, host, 81
- MalumpBa2012: distribution, economic importance, 30,41
- MalumpKa2011a: distribution, host, illustration, 54,57
- MamukYuCa2024: chemical control, natural enemies,
- Mangou2006: biological control, distribution, host,
- MangouAli2009: biological control, natural enemies,
- Mani1994b: biological control, distribution, host, 351-355
- ManiKr1990d: biological control, 122-123
- ManiKr1994: biological control, distribution, economic importance, host, 12-13
- ManiKr1997c: biological control, distribution, host, 45-47
- ManiKr2000a: biological control, 187-189
- MansooShRa2024: chemical control, resistance,
- MansouBeSu2018: chemical control, economic importance, 1-20
- MansouGrSu2016: biological control, diagnosis, economic importance, host, natural enemies, 5-7
- MansouKaLa2009: diagnosis, distribution, host, life history, male, natural enemies,
- MansouMaLa2011: ant association, distribution, host, 197
- MardinSaMi2025: distribution, phylogeny, 201, 203
- Marott1987a: distribution, host, 110
- MarottTr1995a: taxonomy, 70-71
- Martin1954: distribution, economic importance, host, 113-116
- Martin1986: distribution, host, taxonomy, 96
- MartinClZa2024: description, distribution, host, illustration, survey, 157, 164-165
- MartinGaRi2003: biological control, distribution, host, life history, 149-161
- MartinGaRi2003a: biological control, 139-147
- MartinLa2011: catalog, distribution, host, 48
- MartinMoGa2008: control, economic importance, male, phenology,
- Martor1976: distribution, host, 11-248
- Masten2007: distribution, host, taxonomy, 1-242
- MastenSiKr2008: distribution, host, 157-168
- Matile1976a: economic importance, 2
- Matile1978: distribution, host, 52
- Matile1984c: distribution, host, 227
- MatileEt2006: distribution, host, 182
- MatileGePi2004: distribution, host, 35-37
- MatileOr2001: distribution, host, 195
- MazzeoSuRu2008: distribution, host, 149-152
- McCallMiHo2022: ant association, biological control,
- McCoy1985: biological control, 481-499
- McKenz1967: description, distribution, host, illustration, taxonomy, 280-283
- McLeod1939: biological control, distribution, economic importance, host, 62-68
- McLeod1962: biological control, distribution, economic importance, host, 1-33
- MdelleAdZo2019: distribution, economic importance, host, illustration, phenology,
- MealybSt1987: control, economic importance, life history, 24-30
- Melis1930: distribution, host, 14
- MendelBlIs1991: chemical control, 103-112
- MendelBlWy1992: biological control, distribution, host, 45S-49S
- MendelBlZe1992: chemistry, ecology, life history, 118-124
- MendelGrBl1995: biological control, 55-64
- MendelGrSt2001: biological control, distribution, economic importance, host, 251-265
- MendelJaPr2012: behavior,
- MendelPrJa2012: biological control, ecology, 395-394
- MesselViLe2016: biological control, natural enemies,
- MessenBiVa1976: biological control, 543-563
- MetcalMe1993: control, distribution, economic importance, host,
- Meurge2011: distribution, 82
- Meyerd1983: biological control, distribution, host, 1275-1277
- MeyerdChSu1981: distribution, ecology, host, life history, 662-664
- MeyerdFrCh1981: chemical control, economic importance, 49-52
- MeyerdFrHa1979: biological control, chemical control, 893-895
- MeyerdFrHa1982: biological control, chemical control, 134-136
- MeyerdHaDe1978: biological control, 295-299
- MichelHa1995: biological control, chemical control, distribution, economic importance, host, 277-284
- MifsudMaRu2014: distribution, host, 515
- Millar2002: illustration, 229
- MilleHeCa2016: distribution, economic importance, host, natural enemies, 151
- Mills1984: economic importance, 142-144
- MilonaKoKo2008: distribution, host, 33
- MilonaKoMa2011: biological control, ecology, natural enemies,
- MirabaGaCa2018: distribution, host, 194
- ModaffGiUr2025: biological control, chemical control, chemistry, ecology,
- ModaffRiMo2024: chemistry, control,
- ModaffUrAu2025: biological control, 2
- Moghad2009: distribution, host, 24-25
- Moghad2013: description, distribution, host, illustration, structure, taxonomy, 5, 63-65, 99
- Moghad2013a: distribution, host, 70-71
- Moghad2020: distribution, key, 384,e388
- MoghadAbFa2021: control, diagnosis, distribution, economic importance, illustration, life history, natural enemies, 287-289
- MoghadEs2014: key, 59
- MoghadWa2022: diagnosis, distribution, host, key, natural enemies, taxonomy, 11, 107-109, 147
- MoghadWa2024: economic importance, 18
- MohammAbRa2025: economic importance, 264
- MohammGh2008: distribution, 155
- MoharuBa2015: distribution, host, 102-103
- Moir2020: climate change, economic importance, 1
- Monast1958: control, economic importance, 131-165
- MonastZa1959: biological control, description, distribution, economic importance, host, 27-49
- MondalTaAh2025: anatomy, chemical control, illustration,
- MongueMaGr2024: genetic structure, genome, 3-5
- MongueWoMa2025: evolution, genome, male, reproduction,
- MonteiWoPe2019: distribution, host, 393
- Moore1988: biological control, 212-213, 216
- MorandPaGr2015: biology, distribution, host,
- MorenoFaEw1984: behavior, life history, structure, 32-38
- MorrisKi1977: biological control, 183-217
- MullenScNo2016: DNA sequencing, genetics, phylogeny, 209
- Muma1969: biological control, 863-870
- MumaSeDe1961: biological control, 1-39
- MuniapSuSe2026: biological control, distribution, ecology, history, host, natural enemies, 6
- Murray1978: biological control, distribution, host, 139-142
- Murray1978a: biological control, chemical control, distribution, host, 143-147
- Murray1984: biological control, distribution, host, 141-146
- MurthyChRa2024a: DNA, 341
- MurthyChRa2025: distribution, host, illustration, survey, 19
- MustuDeKa2015: biological control, entomopathogenic fungi, 2
- MustuKiUl2008: biological control, 360-367
- NGuessYaNg2019: distribution, host, 3, 4
- NRC1969: biological control, chemical control, ecology, economic importance, taxonomy,
- NamadaUtRa2025: control, entomopathogenic fungi, host, vector, 3, 13, 16
- Nath1972: distribution, host, 6
- NavarrBeAc2025: DNA, diagnosis, male, molecular biology, nymph,
- Nelson1960: behavior, life history, structure, 111-137
- Nelson1961: life history, structure, 85-91
- NestelCoSa1995: ecology, life history, 506-512
- New1975: biological control, 115
- Newste1908: distribution, host, 7
- Newton1953: disease transmission, host, 40
- NhuQuCh2025: biology, ecology, heat-treatment, natural enemies,
- Nicol1952: chemical control, 120
- NiekerMa2012: biological control,
- NiekerMa2013: biological control,
- NormarNoCa2025: phylogeny, 3
- NormarOkMo2019: distribution, host, phylogeny, taxonomy, 10,36
- NukmalRoAp2017: control, natural enemies,
- Nur1966: structure, 439-448
- Nur1967: structure, 376-389
- Nur1968: structure, 202-209
- ObodjiNGNG2015: virus, virus transmission,
- ObokWeAl2018: virus transmission,
- OdinokKuIs1991: chemistry, 555-558
- Olitsk1925: life history, 442
- OliveiSaOl2023: DNA, phylogeny,
- OliverGaWo2003: biological control, distribution, economic importance, host, life history, 163-168
- OmerCoJoWh1946: biological control, 154
- Oncuer1994: distribution, economic importance, host,
- OrlinsRzSh1989: biological control, 25-26
- OrtuDe1982: life history, structure, 341-353
- Osborn1898: distribution, host, 225
- OuvrarKoGu2013: biological control, host, illustration, physiology, 2
- OzdemirYu2024: bacteria, biological control, natural enemies,
- OzgokcKaKi2021: life history,
- OzkanGuUy2001: biological control, distribution, host, 135-166
- PachecBeBl2014: distribution, economic importance, host, life history, phylogeny, 1-13
- PachecBoPr2016: GLRV, biology, control, diagnosis, distribution, illustration, natural enemies,
- PachecKaBa2020: distribution, host, key, phylogeny, 505, 506, 514
- PachecKaMa2017: DNA sequencing, distribution, host, 3
- PachecKaSi2019: distribution, host, phylogeny, 531, 535
- PadillMaNa2016: distribution, host, 558
- Paik1972: distribution, host, 1-4
- PalmaJBl2015: SEM,
- PalmaJBlGu2019: distribution, host, virus transmission, 290
- ParkLeHa2010: distribution, economic importance, host,
- ParrelHaVa1999: biological control, ecology, 819-839
- ParsaKoWi2012: taxonomy, 9
- Passar1989: chemical ecology, chemistry, 1-213
- PassarWe2004: chemical ecology, chemistry, 2896-2899
- PatilPaPa2011: biological control, 693
- PatraGh1994: chemical ecology, chemistry, 1663-1668
- PaulaSoSo2022: natural enemies,
- PavaSiFa2025: chemistry, natural enemies, wax,
- PelegBa1995: distribution, economic importance, host, 263
- Peleka1962: distribution, host, 60
- Peleka1974: biological control, 14-20
- Pelliz2011: distribution, 313
- PellizChMi2015: disease transmission, 72
- PellizFo1996: distribution, 120
- PellizPoSe2011: distribution, host, 293,296
- PerezG2008: distribution, 216
- PerezPeTe2021: ant association, biological control,
- PicartMa2000: distribution, host, 44-46
- PieterMuVu2010: molecular data, taxonomy, 23-28
- PietriBiCo1969: chemical control, 909-915
- PinheiCoCa2023: biological control,
- PinheiMoVi2023: natural enemies,
- PizarrAlBa2020: host, 28
- PlantPrRe1973: chemical control,
- PlataGoBe2023a: ant association,
- PlataGoBe2023c: climate change, distribution, host,
- PlattStMa2018: biological control, 209
- PlattStMa2020: biological control, host,
- PolatUlKa2008: biology, ecology, host, life history, 177-184
- PooranAnMo2026: molecular biology, taxonomy, 9
- PooranAnTh2024: virus transmission, 526
- PoornaSiMu2025: control, distribution, host, 13, 15
- Posnet1947a: distribution, economic importance, host, 388-402
- Poutie1922: biological control, 3-28
- PrakasBh1985: biological control, 29-32
- PrakasKu1985: biological control, distribution, host, 53-54
- Pratt1958: distribution, illustration, taxonomy,
- QinWuQi2011: life history, 1127
- QuezadCoDi1972: distribution, host, 24-25
- Quilis1929: biological control, distribution, host, life history, 73-80
- RacitiBaCo2001: biological control, distribution, host, 49-54
- RacitiTuMa1996: biological control, distribution, host, 652-658
- RadonjHr2025: diagnosis, 158
- RahimOh2019: distribution, 49
- Rajabp2024: natural enemies, 6
- Ramakr1921a: catalog, distribution, host, 343
- Ramakr1941: distribution, host, 107-113
- RamezaKo2026: behavior, host, 4
- RangaSSwPa1959: chemical control, distribution, host, 356-358
- RauppHoSa2001: distribution, host, 203
- Reboul1976: control, distribution, economic importance, host,
- ReddyBh1993: biological control, distribution, host, 15-18
- ReddyPrBh1992: biological control, distribution, 37-44
- ReddyPrKu1988: biological control, distribution, 73-77
- ReddyRaSe2024: chemical control, physiology, 6
- ReddySrBh1991: biological control, life history, 149-150
- Remill1988: distribution, host, 71-72
- RenAsHu2017: DNA, phylogeny, 5, 6
- RidgwaKiCa1977: biological control, 379-416
- RiehlGaLa1964: chemical control, 522-525
- RileyHo1889a: taxonomy, 118
- RipaLuLa2008: biological control, description, distribution, economic importance, host, life history, 192-197
- RipollGa1989: biological control, 399-412
- RisWaRu2010: molecular data, taxonomy, 39-43
- Risbec1949: biological control, distribution, host, 578-581
- Risso1813: description, distribution, host, taxonomy, 416-420
- Rivnay1945: biological control, 117-122
- Rivnay1961: distribution, economic importance, host, life history, 119-124
- RizzoMoCa2024: DNA, 44
- RodrigMaTe2025: natural enemies,
- RodrigSiGo2021: ecology, host, life history,
- RosadeMo2021: distribution, host,
- RoscigCa1985: economic importance, 186-188
- Rosen1969: biological control, 45-53
- RosenDe1973: biological control, taxonomy, 215-222
- RosenbSaSa2012: ecology, molecular data, physiology, 2357-2368
- RossHaOk2012: phylogeny, taxonomy, 199
- RossMoDe2024: biology, genome, illustration, reproduction, taxonomy,
- RossPeSh2010a: life history, 54-65
- RossShPe2008: molecular data, 37
- RostamHuSh2024: biological control, natural enemies, 1408, 1410-1414
- RothwaClWi2004: chemical control, 1239-1244
- Rotund1975a: structure, 3-9
- RotundTr1974: structure, 121-131
- RotundTr1975a: behavior, biological control, chemistry, life history, structure, 172-179
- RotundTr1976a: structure, 99-107
- RotundTr1980: structure, 431-435
- RotundTr1980a: structure, 167-170
- RotundTr1982: structure, 475-478
- RotundTr1983: structure, 97-101
- Rubin1985: biological control, 530-533
- Rubtso1947: biological control, distribution, host, 63-84
- RungMiSc2009: molecular data, taxonomy, 8-12
- RungScEv2008: molecular data, taxonomy, 74
- RungScEv2008a: molecular data, taxonomy, 525-532
- Rungs1970: distribution, economic importance, host, 91-94
- Russo1958: biological control, distribution, 141-147
- Russo1959: chemical control, economic importance, 14-17
- RussoMa1997: distribution, 51
- SaboorHoHa2003: biological control, ecology, life history, 49-54
- SaccagKrPi2008: chemistry, molecular data, taxonomy, 27-33
- SaccoPaD2000: biological control, distribution, host, 3-8
- SadofNeCl2003: ecology, host, life history, 463-469
- SagarrViSt2001a: biological control, 112-116
- Sailer1983: distribution, economic importance, 15-38
- Salama1971: life history, physiology, 1294
- Salaza1999: control, 47
- SancheCaSi2001: biological control, 1-8
- SancheMoAm2026: chemical, control, diagnosis, 4
- SantaCBaSo2009: distribution, host, life history, 13-15
- SantaCPrMa2008: biology, life cycle, nymph,
- SantaCSi2020: ecology,
- SantosCaPe2017: natural enemies,
- Saraiv1939: distribution, host, 103
- SarkarSuUr2014: biological control,
- Sarkie2021: DNA, evolution, genetic structure, morphology, reproduction,
- Savesc1982: description, distribution, host, life history, taxonomy, 261-264
- SayedSoSa1962: life history, taxonomy, 449-457
- SayedSoSa1962a: distribution, host, life history, 459-463
- ScheelBeMe1958: life history, physiology, 303-308
- Schmut1964: distribution, host, 105
- Schmut1969: control, distribution, economic importance, host, taxonomy, 102-104
- Schmut1990: distribution, host,
- Schmut1995: chemical control,
- SchmutPiKl1978: distribution, economic importance, host, 328
- SchneiPo2026: distribution, economic importance, list, 28
- SchutzGoGu2026: taxonomy, 13
- Seabra1921: distribution, host, 96
- Seabra1930: distribution, host, 127
- SeabraBrZi2013: reproduction,
- Seljak2010: distribution, host, 101
- SenaToOl2025: natural enemies,
- ShanLiLi2026: life cycle, 7
- ShiLi1991: distribution, host, 161
- ShrewsBeLe2004: distribution, ecology, host, life history, 425-434
- Signor1869: catalog, taxonomy, 849, 873
- Signor1875c: description, distribution, host, taxonomy, 309-311
- SilvaBrMe2013: behavior,
- SilvaFrCa2008: life history, reproduction, 275
- SilvaFrMe2008: life history, physiology, 274-275
- SilvaMe1996: distribution, economic importance, host, 712
- SilvaMe1999: ecology, economic importance, life history, 89-98
- SilvaMe2001: life history, structure, 347-350
- SilvaMeFr2010: life history, reproduction, 19-21
- SilvaMoBr2018: behavior, reproduction,
- SilvaSSoCo2016: natural enemies, 124, 128
- Silves1921: distribution, economic importance, host, 1-11
- Simant1962a: economic importance, 105-112
- Simant1969: biological control, economic importance, 889-896
- Simmon1958b: biological control, distribution, host, 475-478
- Simmon1969a: biological control, 765-767
- SinghKaPr2025: biological control, 1, 3
- SinhaKuKu2021: control, distribution, host, illustration,
- SiriseWaHe2013: diagnosis, distribution, host, illustration, key, 72,75,77,79
- SismanUl2010: distribution, host, 219-224
- SkelleBuAl2025: diagnosis, 45
- Smetni1991: chemistry, 92-129
- Smit1934: distribution, economic importance, host, 439-441
- SmitBi1934: biological control, chemical control, distribution, economic importance, host, 1-41
- Smith1915: biological control, 525-527
- Smith1917: biological control, 262-268
- Smith1917a: biological control, distribution, host, 108-114
- Smith1958: chemical control, distribution, host, 335-338
- SmithBeBr1997: biological control, distribution, economic importance, host, life history, 82-85
- SmithFl1950: biological control, 362, 376, 378
- SmithFrPa1996: biological control, 57-64
- SmithVa1967: chemical control, 295
- SoaresCaFr2021: natural enemies, 113
- SobiroIbIs2026: Grapevine leafroll disease transmission, biology, distribution, economic importance, host, illustration, 158
- SodanoOuSc2023: ant association, bacteria, morphology, 5
- SorokeNeBa2003: biological control, chemistry, 163-168
- SosanAjUd2020: distribution, ecology, host,
- SoyluOr1977: biological control, 77-112
- Speare1922: biological control, 1-18
- Staedl1976: life history, physiology, 228
- StathaEcBa2023: control, economic importance, 2
- Stimme1987: distribution, host, 24-25
- StoianGrTo2025: chromosome, evolution, genome,
- SuSu1996: biological control, 714
- Suh2019a: key, 5
- Suh2019b: natural enemies, 3
- Suh2020: distribution, host, 3-7, 9-21
- SumaRuDu2002: chemical ecology, chemistry, life history, 305-312
- SumaRuFr2014: control,
- SureshKuGe2005a: biological control, chemical control, 360
- Sweetm1958: biological control, economic importance, 449-458
- SwinglWe1896: distribution, host, 1-5
- Swirsk1985: economic importance, 781-799
- SwirskWyIz2002: biological control, distribution, economic importance, host, life history, taxonomy, 64-68
- SzitaFeBe2017: distribution, host, 473
- SzitaFeGe2023: key, 5
- SzitaGeSi2025: climate change, dispersal, distribution,
- Szulcz1926: distribution, host, 137-143
- TabataIcMo2017: chemistry, reproduction, 8
- TabataLo2002: chemical control, 30
- Tachik1959: biological control, distribution, economic importance, host, 1055-1058
- Talhou1969: distribution, host, 103-105
- Talhou1973: control, natural enemies, 457
- Talhou2002: distribution, economic importance, host, 85-86
- TanakaChKa2025: phylogeny, 580, 591
- Tang1977: description, distribution, host, illustration, taxonomy, 32-34
- Tang1992: description, distribution, host, taxonomy, 367-369
- Tang2001: distribution, 3
- TangHaTa1992: distribution, host, 6
- TangXiLi2025: DNA, behavior, biological control, 2
- Tao1999: distribution, host, taxonomy, 24
- Targio1881: description, distribution, host, taxonomy, 137-139
- Teodor2018: distribution, history, 46
- TerGri1956: biological control, description, distribution, host, taxonomy, 33-35
- TerGri1963: description, distribution, host, illustration, taxonomy, 16, 17
- TerGri1966a: distribution, host, 372
- TerGri1973: description, distribution, host, illustration, taxonomy, 82-85
- Terezn1975: description, distribution, host, illustration, taxonomy, 226-228
- Thaman2018: ant association, 2
- ThaoWiBu2001: chemistry, economic importance, structure, 457-458
- Thomps1958: biological control, distribution, economic importance, host, 479-482
- TirunaSa2024: distribution, natural enemies, record,
- TollerRuDo2004: biological control, distribution, host, 213-217
- TongOmWa2024: genetics, genome, life cycle, male, reproduction, 1
- Topakc2026: distribution, ecology, host, illustration, natural enemies, survey, 6-7, 12
- TrabouBe1965: biological control, distribution, host, 1-13
- Tranfa1978: taxonomy, 91-93
- TranfaTr1982: structure, 1-5
- TremblRoTr1983: structure, taxonomy, 435-437
- TremblTrRo1977: structure, taxonomy, 113-135
- TrenchTo2014: distribution, 67
- TrenchTo2014: distribution, 67
- TrenchTrTo2010: distribution, host, 114-123
- Trjapi1989: biological control, 117, 125, 127, 145,
- Tsalev1968: distribution, host, 206
- Tsalev1972: biological control, distribution, host, 81
- Tschor1939: distribution, host, 89
- TuWuLe1988: description, distribution, host, illustration, taxonomy, 80-81, 92
- Tuncyu1970a: distribution, host, 67-80
- TurgutUl2007: ant association, 313
- TurnbuCh1961: biological control, distribution, host,
- UlgentCa2004: distribution, host, 79-84
- UlgentDo2019: distribution, host, 487
- UlgentErYa2022: distribution, host, S126
- UlubasKaKi2007: molecular data, 232-238
- UlusoyKaBi2022: control, wax secretion,
- UneliuElTw2011: molecular data, 170
- UrbaneRoVa2024: biological control, chemical ecology, natural enemies,
- UsmanPu1955: distribution, host, 47
- UygunSeEr1998: distribution, host, 183-191
- VacantPuRa1996: biological control, 712
- VanHarCoWi1990: distribution, host, 133
- Varshn1992: distribution, host, 57
- VeaFiJa2025: genetics, genome, male, reproduction, 5-6, 12
- VeaGr2015: phylogeny, 64
- VeaGr2016: evolution, 4
- VercheSaEs2025: behavior, control, life cycle, 3, 10-11
- VidyaBh2017: distribution, host, natural enemies, 214
- VidyaBh2017: distribution, host, natural enemies, 214-215
- Vierec1915: biological control, 211
- Viggia1970a: distribution, economic importance, host, 47-55
- Viggia1975: biological control, 52-59
- Viggia1979: biological control, distribution, host, 293-296
- ViggiaCa1971: biological control, distribution, economic importance, host, 535-541
- ViggiaCaBo1972: biological control, chemical control, 88-103
- VonEll2025: distribution, 33
- Vos1930: ecology, life history, physiology, structure, 1-81
- Vos1930: host, life history, 159-168
- WakgarGi2003: biological control, 243-254
- Walker1852: taxonomy, 1081
- WalkerWaEv1974: distribution, host, 11
- WalkerWaEv1974: distribution, host, 13
- WangDaTi2019a: DNA sequencing,
- WangLiRu2023: symbionts, 1
- WangZhTi2018: distribution, host, 2
- WaqasShCh2019: reproduction,
- WardOnBo2024: phylogenetics, 4, 9, 12
- Watson2021: distribution, 526
- WatsonBe1932: biological control, distribution, economic importance, host, 41-42
- WatsonKo2022: biology, diagnosis, distribution, host, illustration, natural enemies, 178-182
- WatsonOoGi1995: distribution, host, 41
- WeintrReMo2017: economic importance, host, 10
- WengHu1988: biological control, distribution, host, life history, 105-108
- Willia1962: distribution, host, 39
- Willia1973: distribution, host, 81-91
- Willia1982b: distribution, host, 442-443
- Willia1985: distribution, host, taxonomy, 272-274
- Willia1986c: host, 55
- Willia1999: distribution, host, 92
- Willia2004a: description, distribution, host, illustration, taxonomy, 630-633
- Willia2005: distribution, host, 168
- Willia2013: distribution, host, 189
- Willia2017a: catalog, list of species, 246, 249
- WilliaBe2009: catalog, taxonomy, 16,46
- WilliaBe2015: taxonomy, 162, 163, 166, 167
- WilliaGr1992: description, distribution, host, illustration, taxonomy, 394-397
- WilliaMa2005a: distribution, host, 147
- WilliaMa2009a: taxonomy, 97-101
- WilliaMa2009b: distribution, host, 120
- WilliaMa2012: distribution, 85, 86
- WilliaMi2010: distribution, host, 47
- WilliaMo2000: distribution, host, taxonomy, 33-34, 39-40
- WilliaPa2012: description, taxonomy, 227
- WilliaWa1988a: description, distribution, host, illustration, taxonomy, 162-164
- WoglumLaLa1947: biological control, 818-820
- WoglumLaLa1947: biological control, chemical control, 818-820
- Wolcot1955: distribution, host, 1-5
- WolkGoDu1986: structure, 347-348
- WongChCh1999: description, distribution, host, taxonomy, 6,44
- Wood1962: biological control, distribution, 8-11
- WuXu2022: history, morphology, nomenclature, 164
- WynneHoCo2025: distribution, list of species, 19
- XiYaLi2024: DNA, control, molecular biology,
- Yang1995JS: biological control, distribution, host, life history, 1-83
- YangLyMa2025: behavior, male, reproduction, 123
- Yasnos2001: biological control, distribution, host, 435-440
- YasnosTaCh2005: biological control, distribution, host, 295-302
- YaylaTuSa2020: biological control, natural enemies,
- YerlikBaKa2023: distribution, host, list, 10-11
- YigitCa1995: biological control, distribution, host, 25-26
- YigitCa1995a: biological control, 1-22
- YigitCaKi1992: biological control, chemical control, distribution, host, 251-263
- YigitCaZa1994: biological control, 137-146
- YigitErKa2005: biological control, 383-387
- YokogaYa2009: genebank, molecular data, phylogeny, 57-66
- YooBaCh2025: chemical control, heat-treatment, resistance,
- YunMoAj2025: bacteria, biological control, distribution, host, illustration, molecular biology, phylogeny, record,
- ZadaDuHa2004: chemical ecology, chemistry, ecology, life history, 361-368
- Zahrad1990b: description, distribution, host, illustration, taxonomy, 29-31
- ZarkanApTu2021: distribution, host, 184
- ZarkanFaAp2024: key, 237, 238
- ZarkanHaRe2025: key, 57
- ZarkanNaDj2022: distribution, 189
- ZchoriBePo2005: endosymbionts, 211-221
- ZettleAdFr2017: host, 33
- ZhangDe2023: distribution, host, key, taxonomy, 79, 80, 85
- ZiegleWo1975: distribution, economic importance, host, 1-6
- Zimmer1948: description, distribution, host, illustration, taxonomy, 176, 261-270
- deJongVa1988: biological control, life history, 1079-1096
- deSilvJoSh2002: disease transmission, life history, 537-545
- vanLen2003a: biological control, distribution, host, 1-18
- vanLen2003b: biological control, distribution, host, 167-179
- vanLenHaKl2003: biological control, distribution, host, 265-303
- vanden2001c: biological control, chemical control, distribution, economic importance, host, life history, 196-208

